Introduction
This chapter describes the brain’s hardware, the physical machinery that determines what a viewer can perceive, feel, think, and want when they watch a piece of content (a video, an ad, any creative piece - “content” throughout this book). Specifically, it identifies thirteen properties of the brain that Khozai’s predictions depend on. If any one of these properties were missing from the brain, Khozai’s predictions would go wrong in a specific and traceable way.
What the reader will learn. The brain’s input channels, its regional and network organization, its chemical systems, its global control mechanisms, its connection to the body, its capacity for self-modification, and its architectural principles. These are not abstract neuroscience, they are the hardware constraints that determine what Khozai can and cannot infer about a viewer’s response.
Why. Khozai measures content properties and predicts brain responses. The brain’s architecture determines what those responses can be. Every dimension of the viewer’s subjective experience that Chapter 4 will identify traces back to a specific piece of the hardware described here. Every prediction that Khozai makes through TRIBE v2 (the brain encoding model that predicts whole-brain activation from the video file alone, no scanner, no subjects) is a prediction about activity across the brain described here, cortical (brain’s outer surface) predictions at higher confidence, subcortical (deeper internal structures) predictions at lower confidence. Without this chapter, the rest of the book floats without a physical foundation.
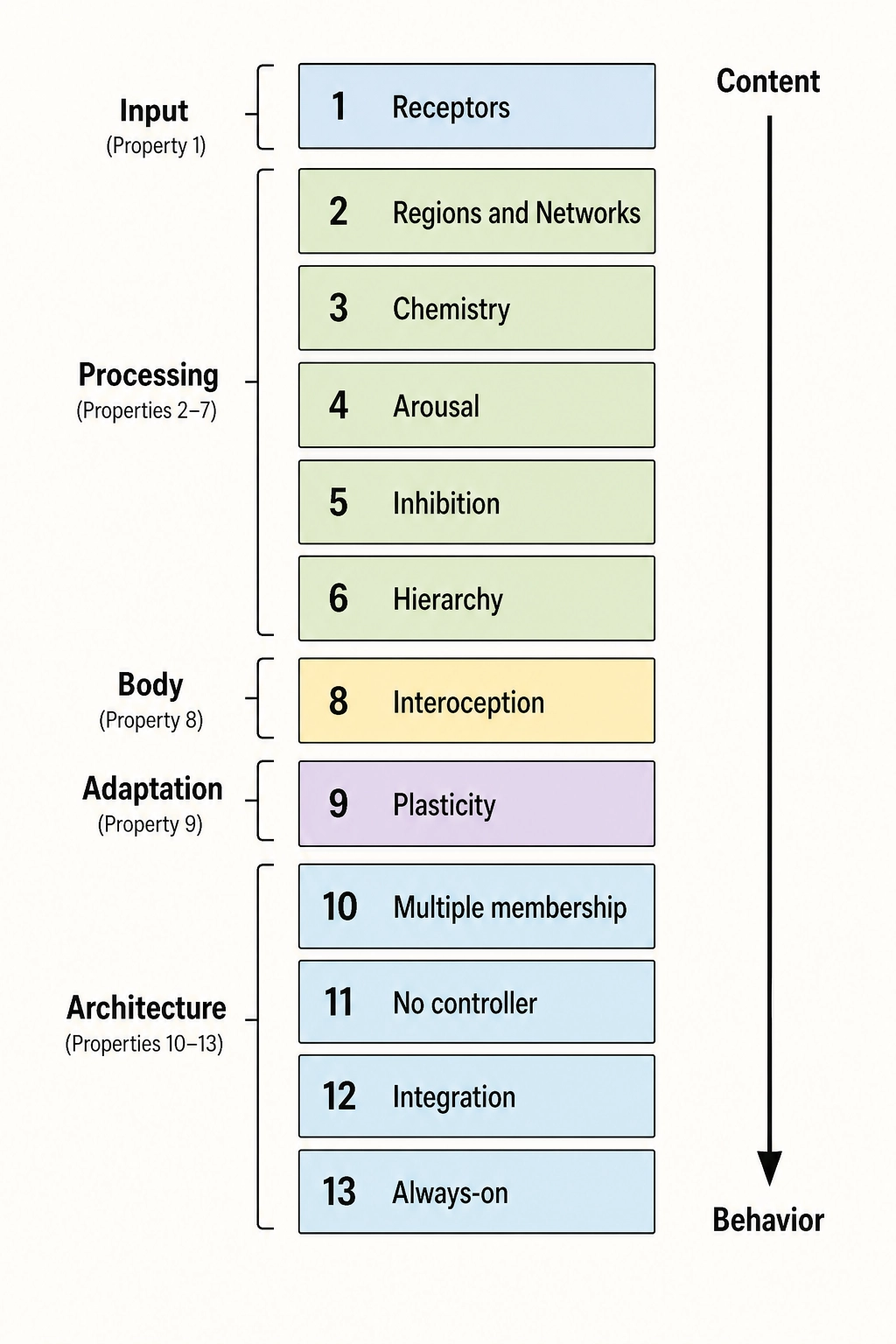
How the chapter is organized. The thirteen properties of the brain are grouped by function: input, processing, chemistry, global controls, body connection and plasticity, and architectural principles. Eight secondary properties follow. The chapter closes by connecting the architecture to Khozai’s framework.
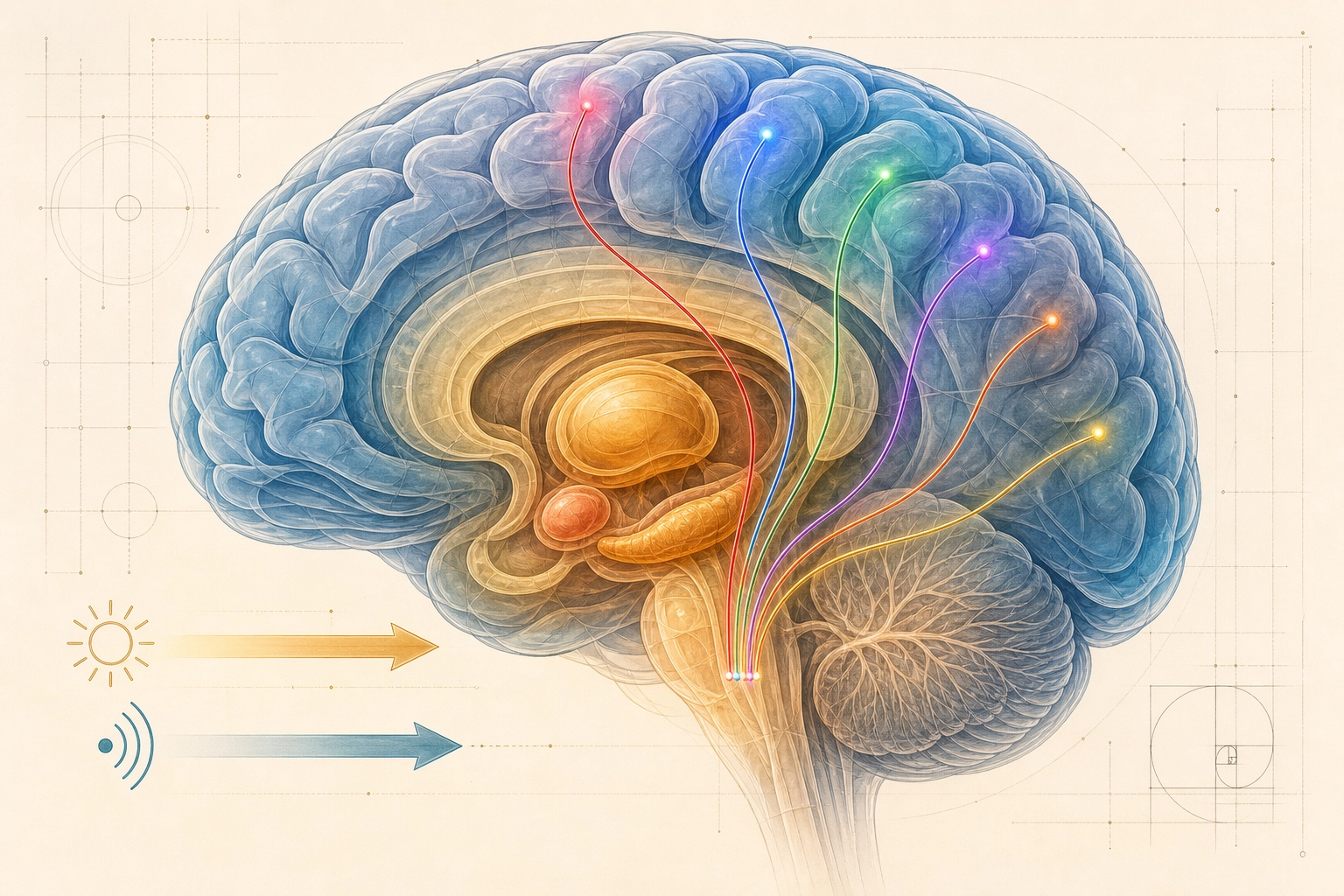
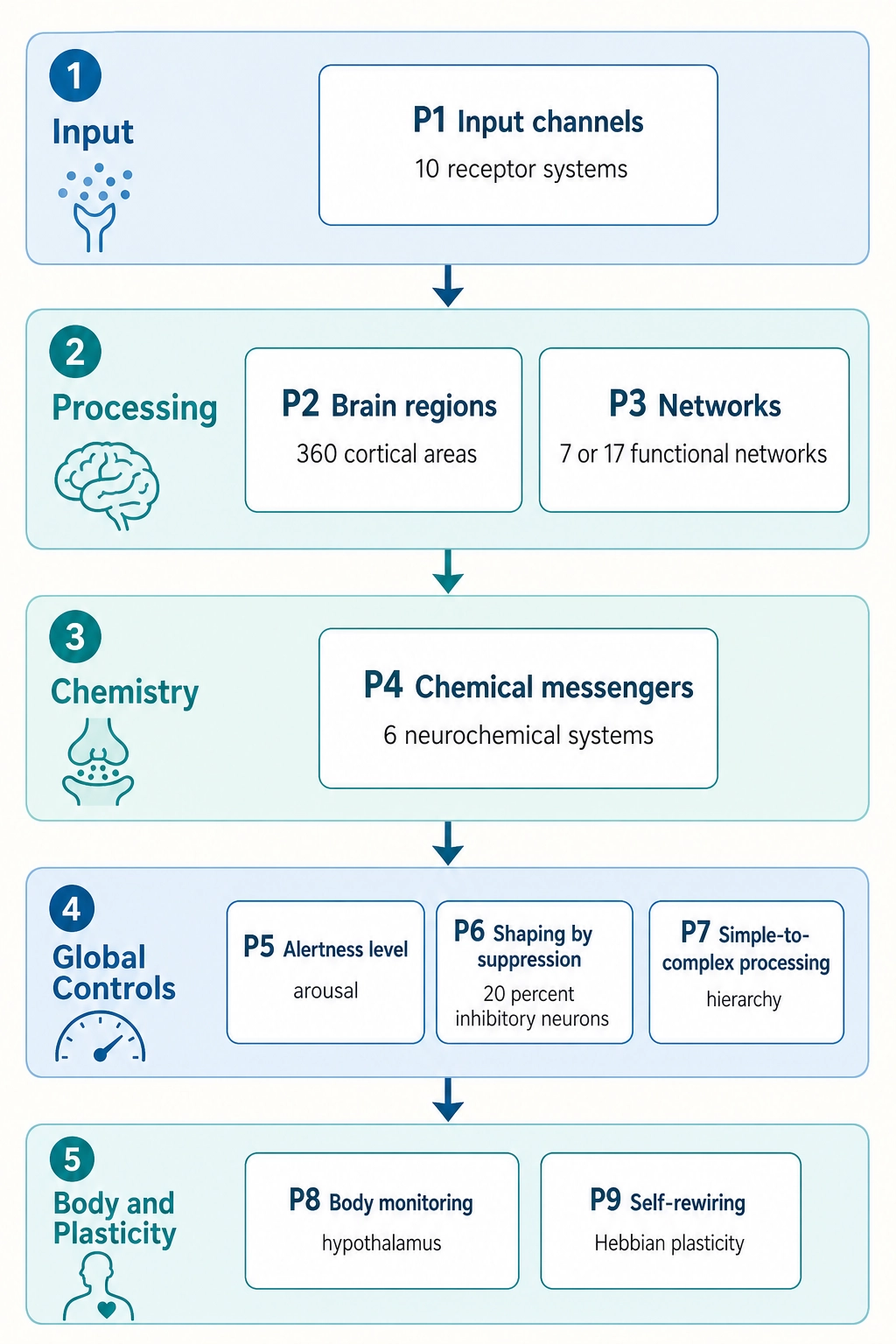
Part 1 - The hardware (Properties 1-9):
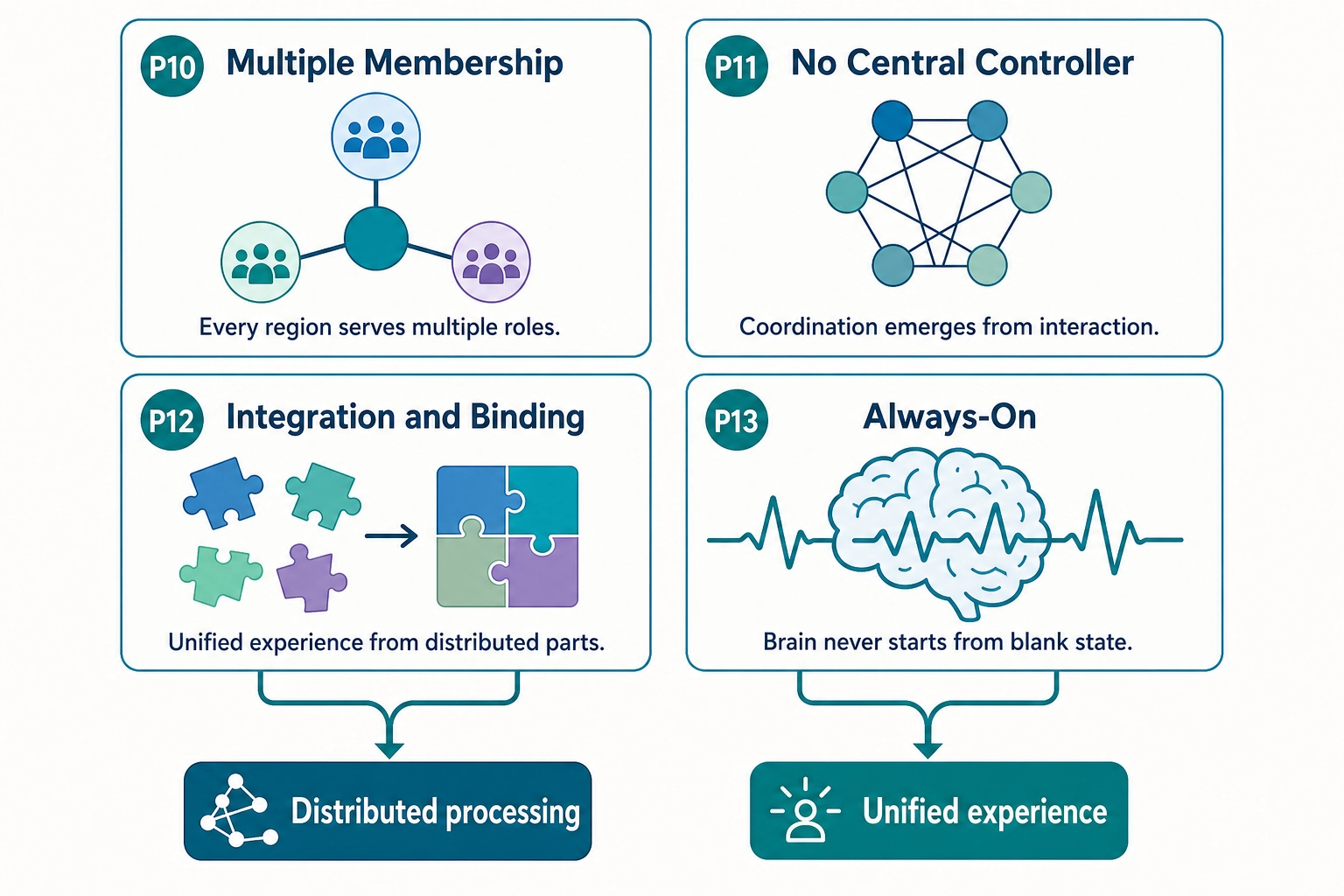
Part 2 - The architectural principles (Properties 10-13):
A terminology note. This chapter inherits Chapter 2’s vocabulary. “Experience” means subjective experience, what the viewer perceives and feels. “Content” means the video or creative piece. In Chapter 2’s vocabulary, a “space” is the complete set of all possible states of something, not a physical location, but a mathematical way of describing every configuration something could take. The four spaces are: Physical Stimulus Space (every possible physical input the brain could receive), Neural State Space (every possible configuration of the brain’s hardware), Experience Space (every possible moment of subjective experience), and Behavioral Output Space (every possible behavioral response). These terms, along with premises, mappings, and reasoning tools from Chapter 2, are used freely in this chapter. Technical brain-anatomy terms that are load-bearing for the argument (fusiform face area, temporal parietal junction, default mode network, primary visual cortex) are glossed in the same sentence on first use. Deep specialist terms that the reader does not need to follow the argument are named but not elaborated - the reader can google them if curious or let them pass without losing the thread.
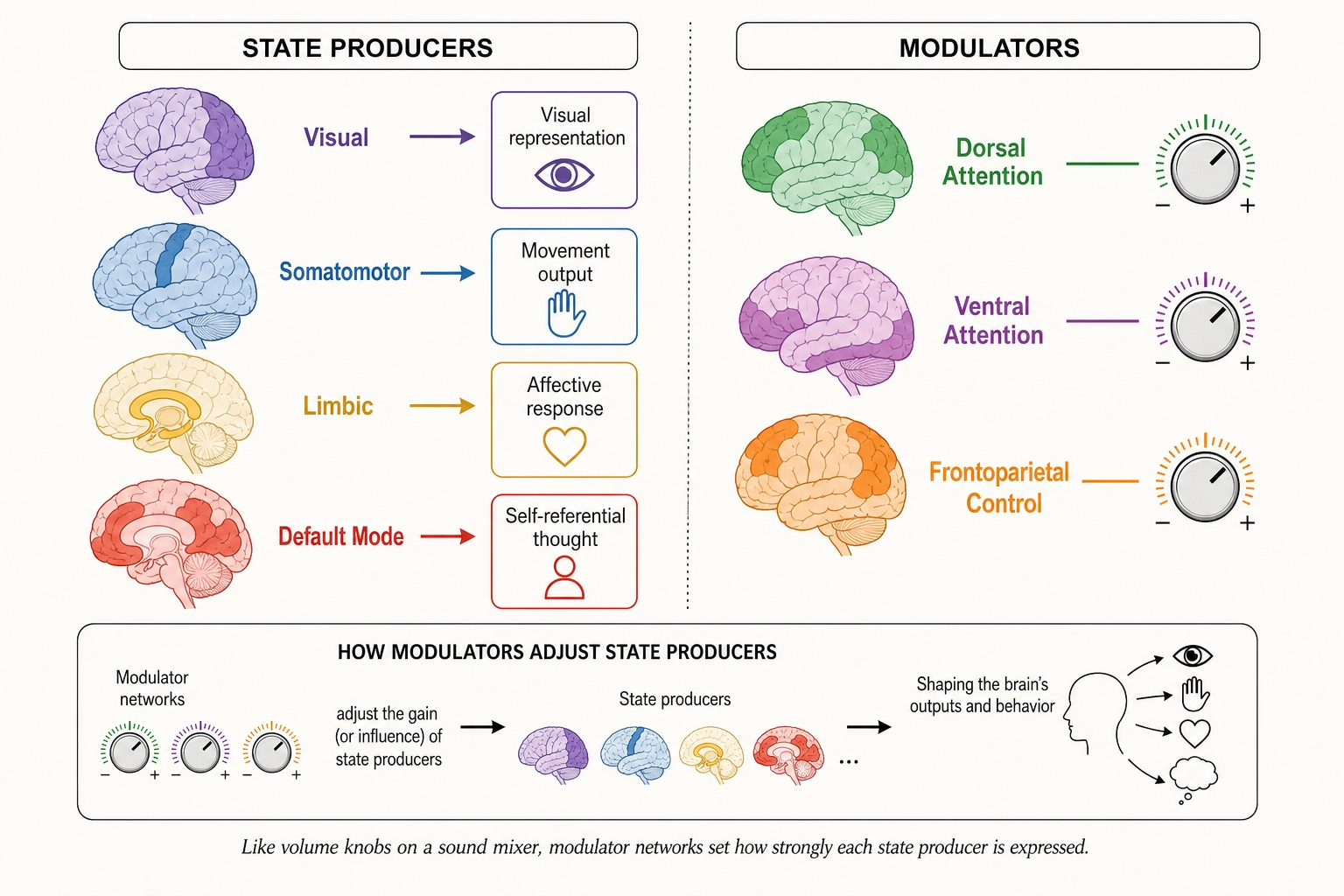
Each brain system introduced in this chapter is classified using the Classification Test (Tool 5 from Chapter 2): destroy or disable the system and observe. Does a specific aspect of subjective experience disappear? The system is a state producer. Do existing aspects change in character but none disappear? The system is a modulator. Does processing efficiency degrade but experiential character remain unchanged? The system is infrastructure, it keeps the machinery running (like electrical wiring in a building) but does not itself produce any specific experience. These classifications appear in the tables throughout the chapter.
1. The Input Interface: Receptors
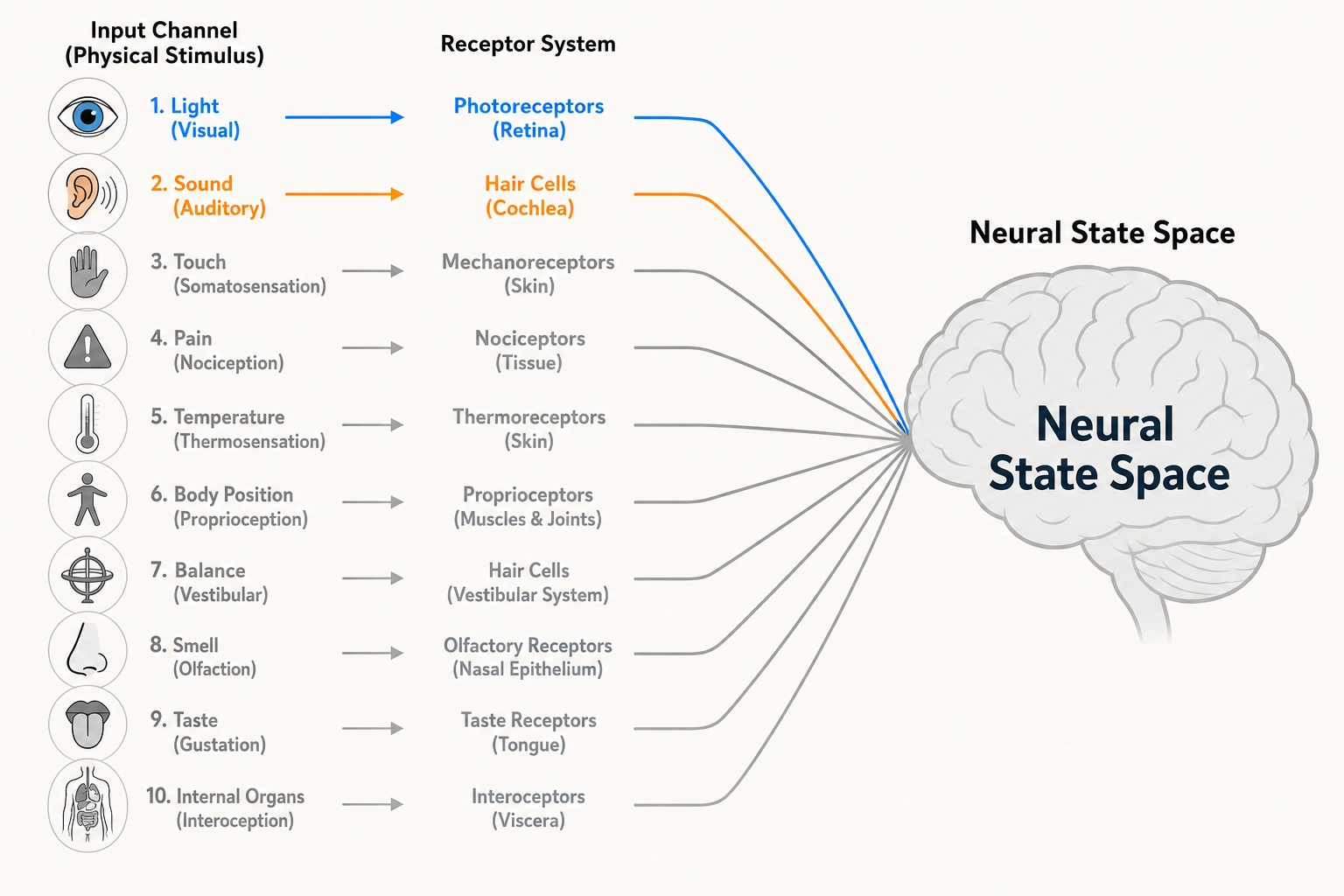
Property 1: The brain has exactly ten receptor systems. They are the only interface between the physical world and the brain.
Video content enters the brain through just two of these ten systems, photoreceptors (vision) and cochlear hair cells (hearing). But the brain was built for all ten, and its internal wiring reflects that full set.
Why does Khozai care about all ten? Two reasons. First, Physical Stimulus Space (the set of all physical energy human receptors can detect) is grounded in Premise 2 and defined by what these receptors can transduce (convert physical energy into neural signals), miss a receptor system, and you miss every experiential dimension that depends on it. Second, the experiential dimensions in Chapter 4 trace back to which receptor systems feed them. The full inventory matters even when only two channels carry the signal.
| # | Receptor System | Physical Dimension | Key Receptor Types | Classification (Tool 5) | What Breaks If Removed |
|---|---|---|---|---|---|
| 1 | Photoreceptors | Light (electromagnetic radiation, 380—700 nm) | Rods, L/M/S cones | State producer | Vision disappears entirely |
| 2 | Cochlear hair cells | Air pressure waves (20—20,000 Hz) | Inner and outer hair cells | State producer | Hearing disappears entirely |
| 3 | Mechanoreceptors | Mechanical deformation of tissue | Meissner, Pacinian, Merkel, Ruffini | State producer | Touch disappears; pain and temperature preserved |
| 4 | Nociceptors | Extreme mechanical force, extreme temperature (>43°C / <15°C), and damage-associated chemicals: bradykinin, prostaglandins, H⁺, K⁺ | A-delta mechanical, A-delta thermal, C polymodal | State producer | Pain disappears entirely |
| 5 | Thermoreceptors | Thermal energy | Warm receptors, cold receptors | State producer | Temperature sensation disappears |
| 6 | Proprioceptors | Muscle/tendon stretch and joint angle | Muscle spindles, Golgi tendon organs, joint receptors | State producer | Body-position awareness disappears (documented in a patient who lost all proprioception below the neck - see section 5) |
| 7 | Vestibular organs | Angular and linear acceleration | 3 semicircular canals, 2 otolith organs | State producer | Balance and spatial orientation disappear |
| 8 | Olfactory receptors | Airborne molecules | ~400 receptor types | State producer | Smell disappears entirely |
| 9 | Gustatory receptors | Dissolved molecules | Sweet / salty / sour / bitter / umami | State producer | Taste disappears entirely |
| 10 | Visceral afferents | Internal organ state | Mechanoreceptors, chemoreceptors, osmoreceptors in organs | State producer | Body-state awareness reduced (documented in patients with insular cortex damage - see section 5) |
Every receptor system is a state producer: destroying it eliminates a specific aspect of subjective experience while leaving all others intact. This is the selective elimination (destroying one structure eliminates one specific aspect of subjective experience while leaving everything else intact) pattern described in Premise 5, the mapping from brain structure (a specific region, receptor system, or circuit) to subjective experience is specific, not diffuse.
No other entry points exist.
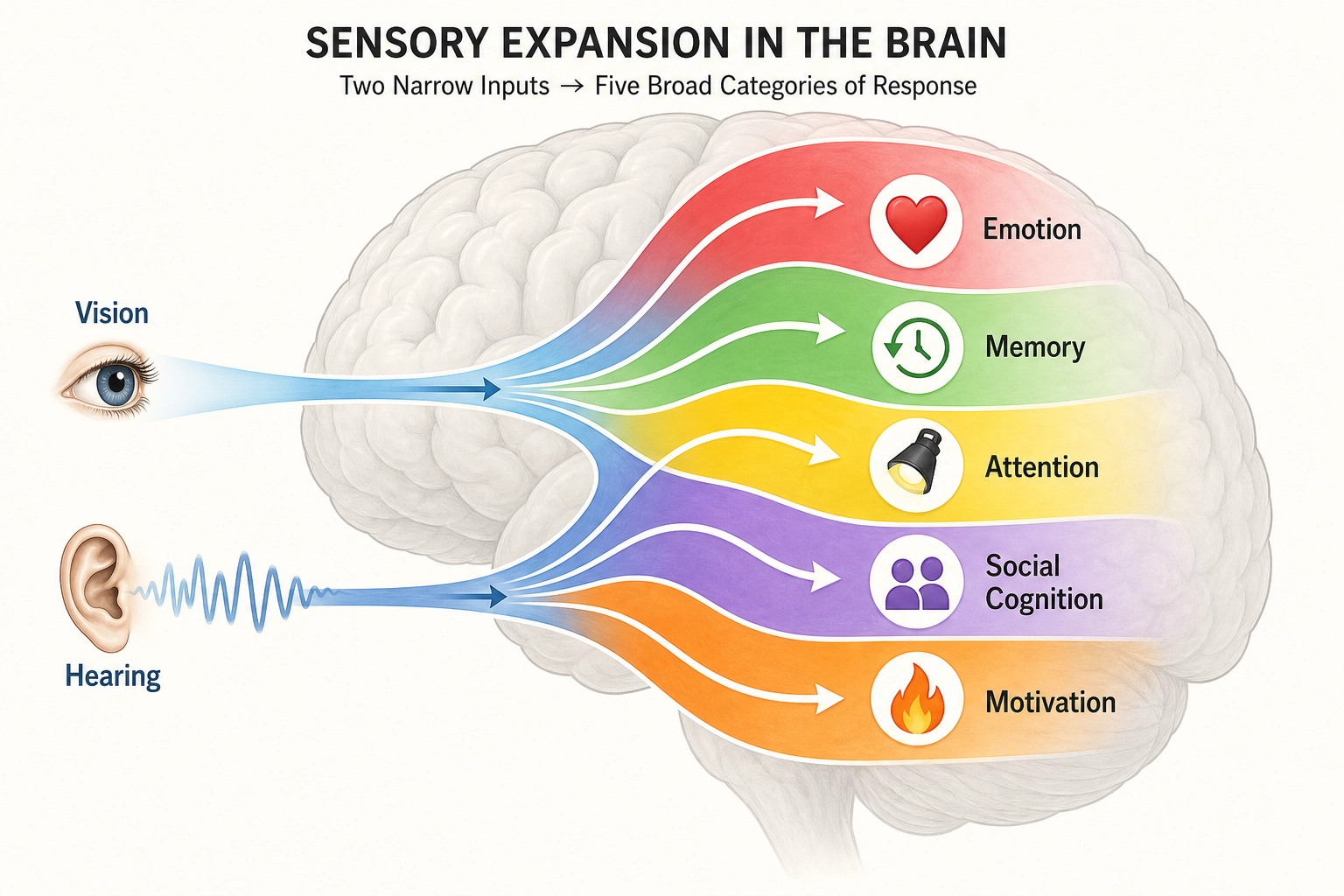
Here is the key asymmetry: video content enters through just two channels, vision and hearing. But the brain’s internal processing does not stay confined to those two channels. Through predictive processing (the brain’s habit of predicting incoming input rather than passively receiving it) and associative networks (connections between brain regions that activate related memories and concepts), a video can activate social cognition, memory, motivation, body-state awareness, and emotion. Two input channels, full experiential activation. This asymmetry is central to Khozai’s design.
Khozai implication. V₀ (the physical-property measurements Khozai extracts from the content file) measures the physical properties that photoreceptors and cochlear hair cells will receive. Physical Stimulus Space (Premise 2) is defined by what all ten receptor systems transduce. Miss a receptor system from the list, and every experiential dimension that depends on it becomes invisible to Khozai’s framework.
Is the list really complete? The balance of evidence strongly favors yes. The Piezo channel discovery [1] (the biophysicist Bertrand Coste and colleagues, Science, 2010) identified the molecular mechanism underlying mechanoreception, it did not add an eleventh receptor system. No candidate for an additional receptor system has emerged.
The receptors define what enters the brain. The next question is what happens to the signal once it gets there.
2. The Processing Architecture: Regions and Networks
2.1. Cortical Organization
Property 2: The brain’s outer surface (the cortex) contains approximately 360 anatomically distinct regions.
Property 3: These cortical regions organize into functional networks, groups of regions whose activity rises and falls together. Different networks operate independently of each other.
The cortex, the wrinkled outer surface of the brain, is where the processing happens that Khozai can predict. It is not a uniform sheet. It is divided into roughly 360 distinct areas, each with its own cell composition, connectivity pattern, and functional role.
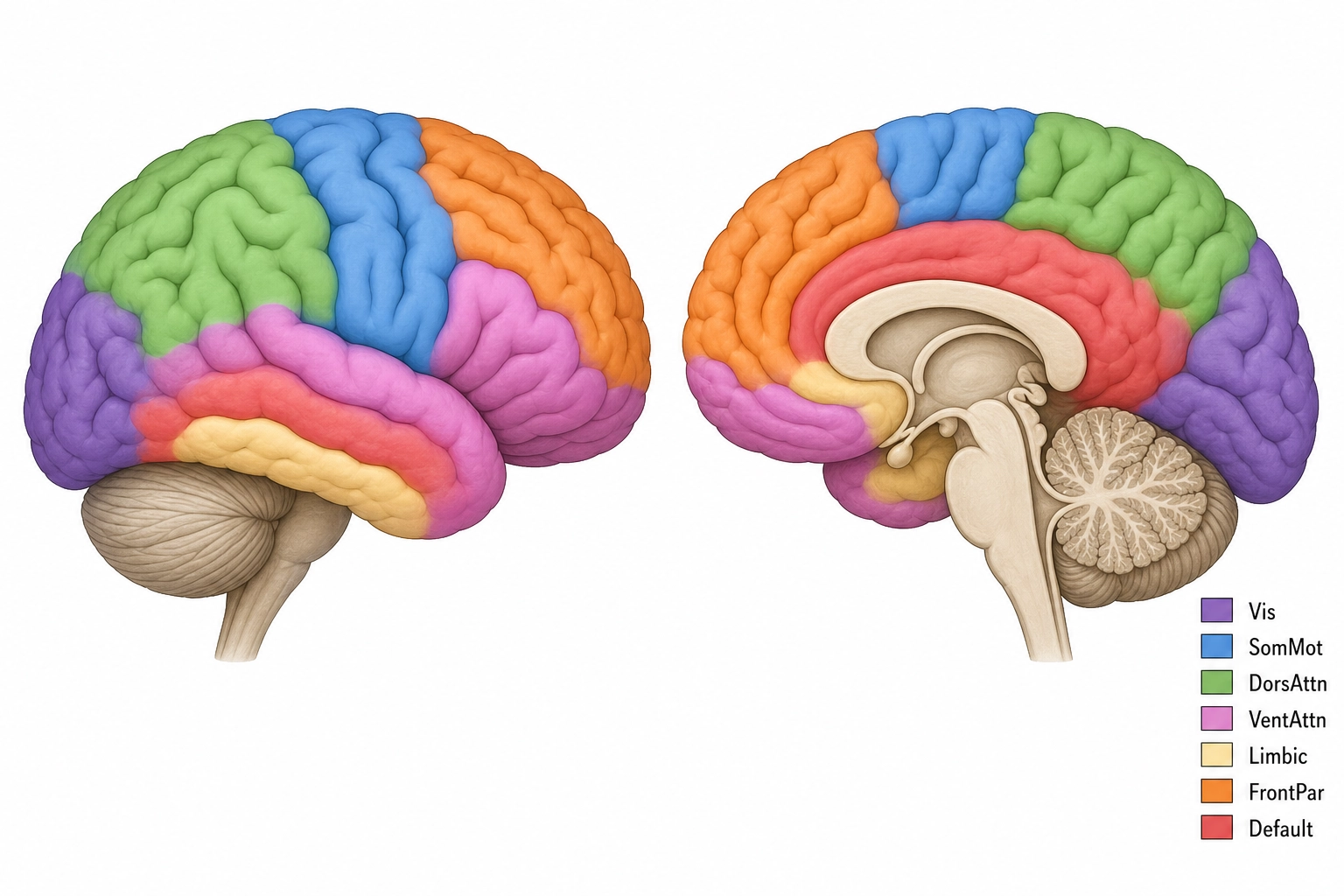
How do we know it is 360? The mapping effort spans over a century. The neuroanatomist Korbinian Brodmann [2] (1909) identified approximately 52 cytoarchitectonic areas by examining cell-layer structure under a microscope. Dr. Matthew Glasser and colleagues [3] (Nature, 2016) refined this to 180 areas per hemisphere, 360 total, using multi-modal MRI data (combining multiple types of brain scans, thickness, myelination, connectivity, and task responses, into one map) from 210 Human Connectome Project subjects. Myelination refers to the fatty insulation coating on nerve fibers. They combined cortical thickness, myelination, functional connectivity, and task activation into a single map. This is the highest-resolution anatomical atlas available.
These 360 regions do not work in isolation. They organize into functional networks, groups of regions whose neural activity rises and falls together. Dr. B.T. Thomas Yeo and colleagues [4] (Journal of Neurophysiology, 2011) analyzed resting-state fMRI (brain scanning done while the person lies still and lets their mind wander, using functional magnetic resonance imaging, which measures blood-flow changes as a proxy for neural activity) data from 1,000 subjects. They found stable network groupings at two resolutions: 7 networks (coarse) and 17 networks (fine).
Khozai uses the seven-network solution as its default resolution (Tool 7, the coarsest resolution at which no operationally relevant information is lost). But all resolutions are available, 7 networks, 17 networks, or the full 360-region atlas. Which resolution Khozai uses for a given analysis depends on what the empirical data shows: if the finer 17-network split produces measurably different behavioral predictions for a given content property, Khozai switches to it. Resolution is a dial, not a fixed setting.
| Network | Core Function | Example Regions | Classification (Tool 5) | Khozai Relevance |
|---|---|---|---|---|
| Visual | Processing visual input | V1 (primary visual cortex, first cortical stage of vision), V2, V4, MT/V5, FFA (fusiform face area, face processing) | State producer | Predicting which visual features the viewer processes |
| Somatomotor | Body sensation and motor control | Primary motor cortex, primary somatosensory cortex, supplementary motor area | State producer | Not directly relevant to video viewing, but activates when the viewer mentally simulates physical actions they see on screen (e.g., watching someone run, dance, or get hit) |
| Dorsal Attention | Top-down, voluntary attention | Intraparietal sulcus, frontal eye fields | Modulator | Predicting where and how strongly the viewer directs attention |
| Ventral Attention / Salience | Bottom-up capture; detecting unexpected events | TPJ (temporal parietal junction, models other people’s intentions), anterior insula, anterior cingulate | State producer / Modulator | Predicting what captures the viewer’s attention involuntarily |
| Limbic | Emotion and memory-related processing | Orbitofrontal cortex, temporal pole, medial temporal regions | State producer | Predicting emotional and memory-related engagement |
| Frontoparietal Control | Flexible, task-dependent control | Lateral prefrontal cortex, inferior parietal lobule | Modulator | Predicting how much mental effort the content demands, e.g., following a complex argument vs. watching a simple visual |
| Default Mode | Self-reference, mental simulation, narrative processing | mPFC (medial prefrontal cortex, self-referential processing), PCC (posterior cingulate cortex), angular gyrus | State producer | Predicting whether the viewer relates content to their own life, imagines future scenarios, or engages in narrative comprehension |
All seven network names match Dr. Yeo’s published labels. The 17-network solution subdivides each of these further, for example, splitting the Visual network into early visual (V1-V3) and higher visual (face, place, object areas). Khozai can operate at either resolution. The choice depends on whether the finer distinctions produce measurably different behavioral outcomes (Tool 7).
2.2. Subcortical Structures
Below the cortex sit older, evolutionarily conserved structures (ancient brain parts shared with other animals, predating the cortex). They handle what the cortex does not: threat detection, reward computation, arousal regulation, body-state monitoring, and motor coordination. Premise 4 establishes that these subcortical structures are finite and complete.
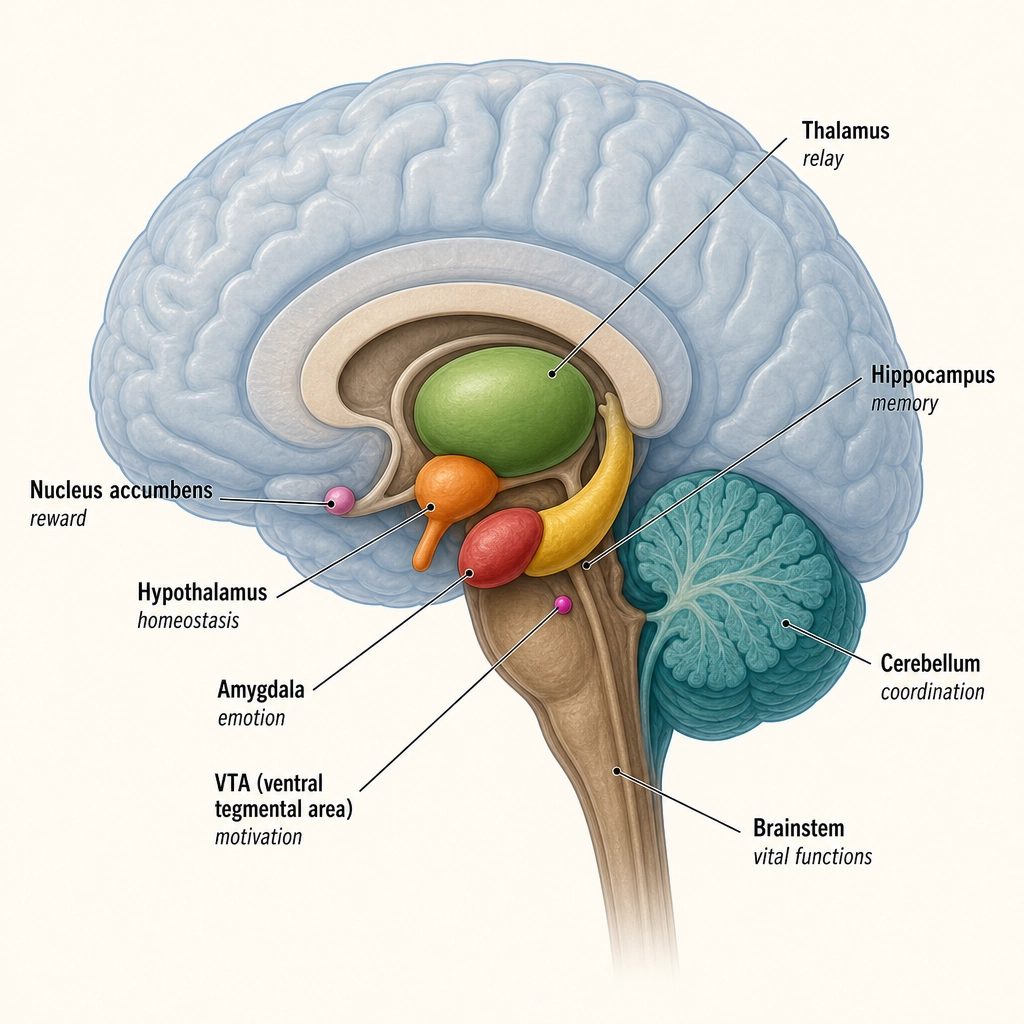
| Subdivision | Key Structures | Primary Functions | Classification (Tool 5) |
|---|---|---|---|
| Diencephalon | Thalamus (relay hub for all senses except smell), hypothalamus (body-state regulation), epithalamus, subthalamus | Sensory relay, homeostasis, circadian rhythm | Infrastructure (thalamus); State producer (hypothalamus) |
| Basal ganglia | Caudate, putamen, globus pallidus, nucleus accumbens (reward), ventral pallidum | Action selection, reward processing, habit formation | State producer (wanting/reward) |
| Limbic subcortical | Amygdala (threat/salience detection), hippocampal formation (memory), BNST, septal nuclei | Fear, memory consolidation, sustained anxiety | State producer |
| Basal forebrain | Nucleus basalis of Meynert, medial septal nucleus, diagonal band of Broca | Acetylcholine supply to cortex; attention and learning modulation | Modulator |
| Brainstem, Midbrain | VTA (dopamine source for wanting), substantia nigra, superior/inferior colliculi, periaqueductal gray, red nucleus, pedunculopontine nucleus | Reward, movement, orienting, pain modulation | State producer (VTA) |
| Brainstem, Pons | Locus coeruleus (norepinephrine source), raphe nuclei, parabrachial nucleus, pontine nuclei | Arousal, alertness, exploration/exploitation | Modulator |
| Brainstem, Medulla | Caudal raphe, nucleus tractus solitarius, RVLM, area postrema, reticular formation, inferior olive | Autonomic regulation, arousal (the reticular activating system spans midbrain through medulla) | Infrastructure / Modulator |
| Cerebellum | Cerebellar cortex, deep cerebellar nuclei | Motor coordination, timing, prediction | Infrastructure |
| Other | Claustrum, pituitary gland | Unclear (claustrum); endocrine output (pituitary) | Infrastructure |
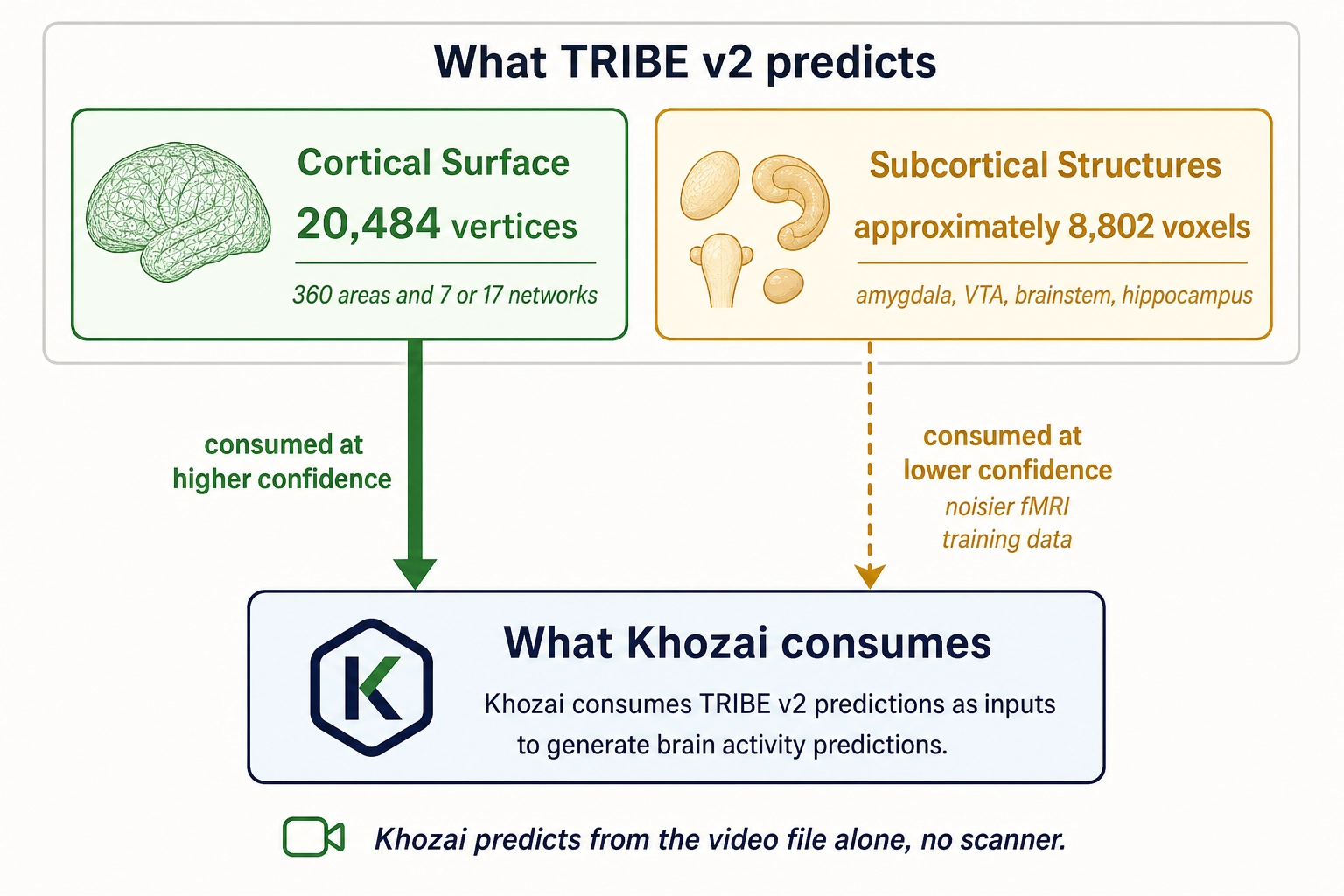
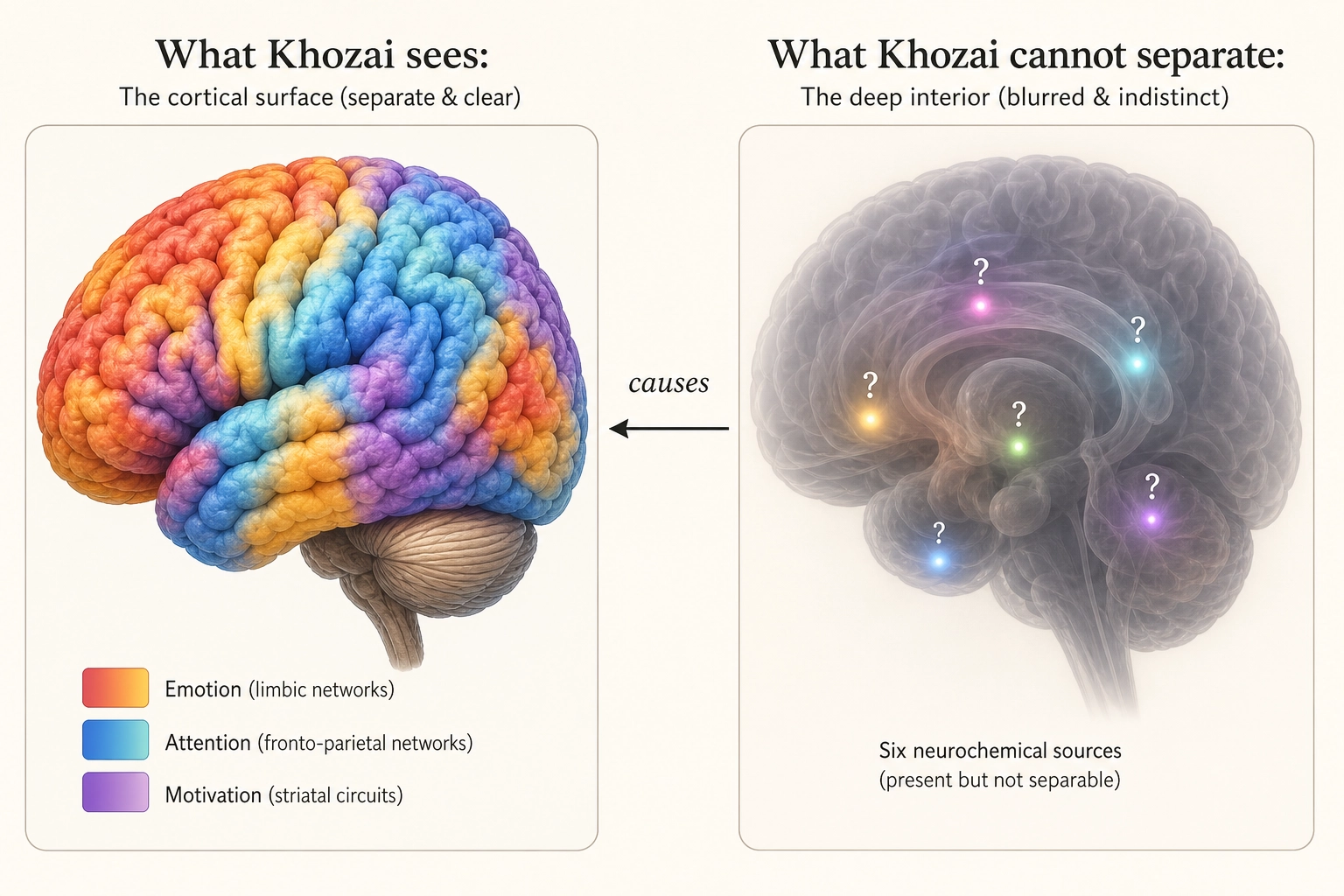
The critical framing for Khozai. Khozai consumes whole-brain predictions from TRIBE v2, both cortical and subcortical, at different confidence tiers.
TRIBE v2 emits whole-brain predictions: 20,484 measurement points on the cortical surface (vertices) plus approximately 8,802 measurement points in subcortical structures (voxels). Khozai consumes both, but at different confidence levels. Here is why the confidence differs.
fMRI scanners work by detecting blood-flow changes near active neurons. The cortex is a large, thin sheet right at the brain’s surface, close to the scanner’s sensors, with a strong and relatively clean signal. Subcortical structures are small and deep inside the brain, farther from the sensors, with a weaker and noisier signal. The difference is physical: a small structure buried deep simply produces less detectable blood-flow change per unit of neural activity than a large surface structure close to the coils. The neuroimaging researcher Max Keuken and colleagues [15] (2018) documented this systematically in Brain Topography, showing that absolute signal-to-noise ratio is substantially lower in subcortical structures due to distance from coil elements, g-factor penalties from parallel imaging, and unfavorable receive profiles.
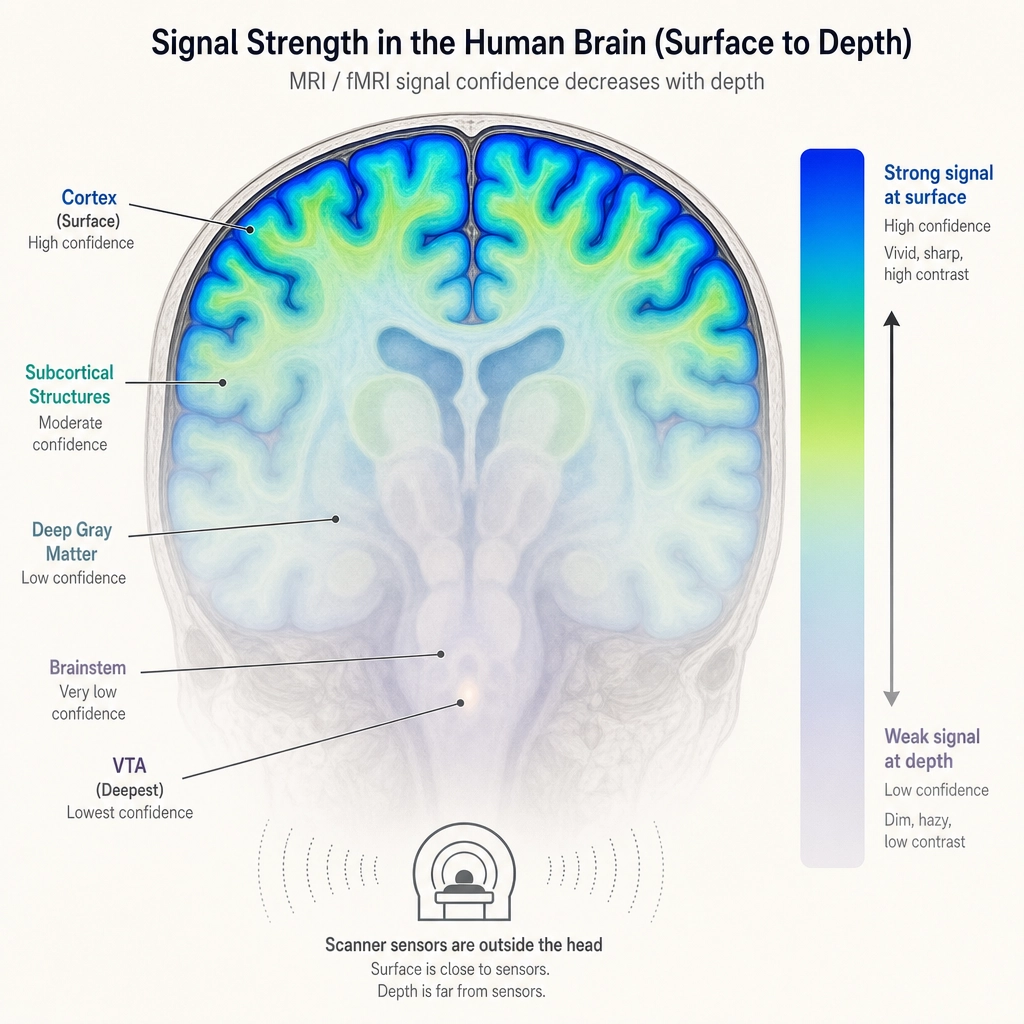
Khozai does not use a scanner, it predicts from the video file alone. But TRIBE v2, the encoding model it uses, was trained on fMRI data, so it inherits this quality gap. Its cortical predictions are trained on cleaner data and produce more accurate reconstructions (where “reconstruction” means: how closely the model’s predicted activation pattern matches what a real scanner would measure). Its subcortical predictions are trained on noisier data and are correspondingly less precise.
Khozai’s response to this is not to discard the noisier signal. It is to tag it. Cortical predictions enter the pipeline at higher initial confidence. Subcortical predictions enter at lower initial confidence. The structures those subcortical predictions represent, reward processing (VTA/dopamine), arousal regulation (brainstem), body-state monitoring (hypothalamus), threat detection (amygdala), memory consolidation (hippocampus), are among the most relevant signals for content engagement. Throwing them away because the measurement is noisier would be like ignoring a witness because their eyesight is imperfect: the testimony is still valuable, it just needs a reliability tag.
The confidence gap is an empirical starting point, not a permanent verdict. The calibration governance framework (Chapter 10) handles graded confidence natively. If Khozai’s experiments eventually show that subcortical predictions carry no useful signal, the confidence will be revised downward. If they carry more signal than expected, the confidence will be revised upward. Khozai learns.
Regions and networks define WHERE the brain processes content. What determines HOW it processes - how motivated, how alert, how emotionally reactive the viewer is - is the chemical environment those regions operate in.
3. The Chemical Systems: Neurochemistry
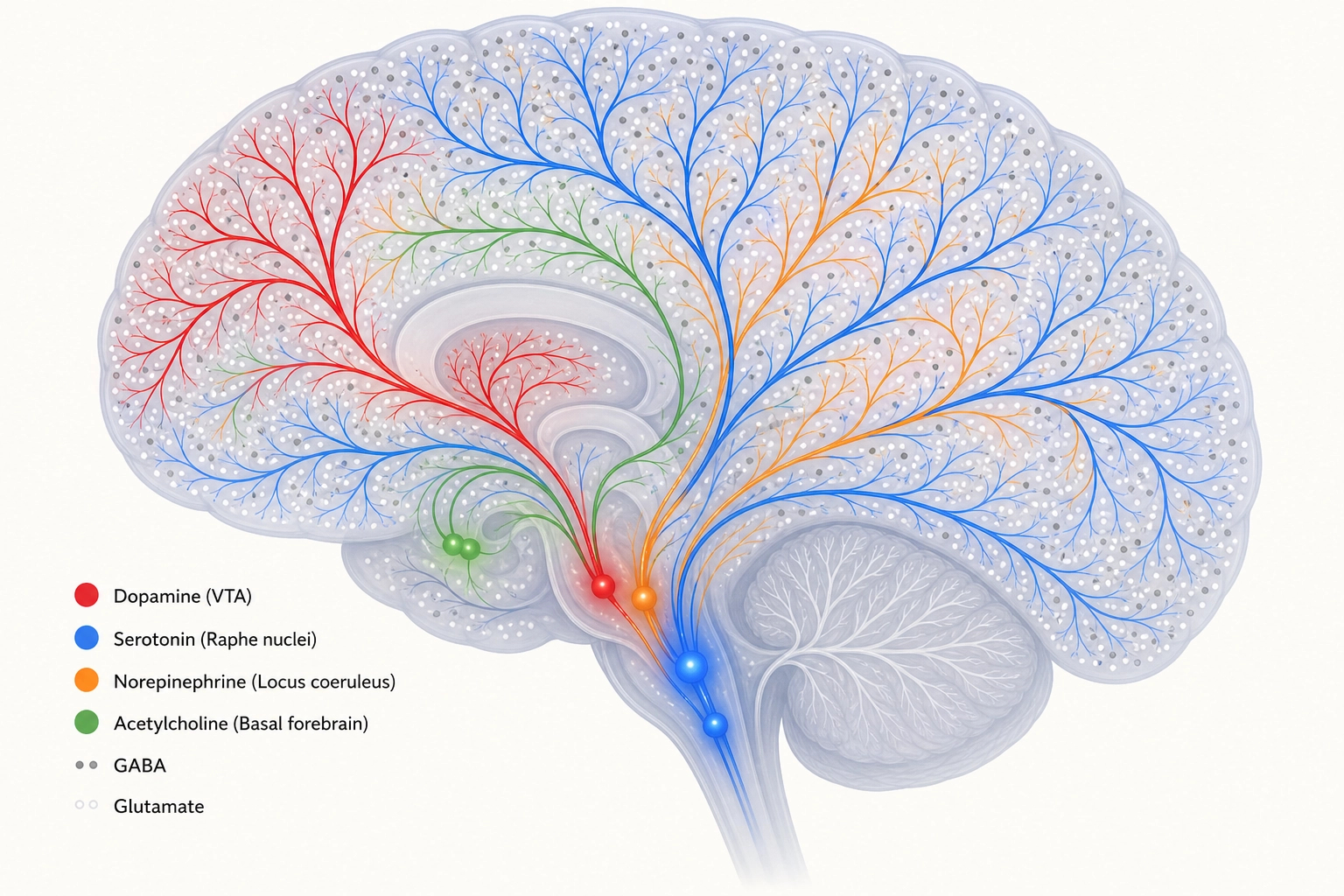
Property 4: Six major chemical messenger systems (neurochemical systems) modulate how the brain processes information.
The brain’s processing mode is not fixed. It is continuously adjusted by chemical messengers, small clusters of neurons deep in the brainstem and basal forebrain that send signals widely across the brain.
Think of these chemical systems as volume knobs and tone controls on the brain’s processing. The cortical regions from Section 2 decide WHAT the brain is seeing, a face, a landscape, a word. The chemical systems decide HOW MUCH the brain cares, how motivated, how alert, how emotionally reactive, how focused. The same video shown to the same viewer at two different dopamine levels produces a different response, not because the content changed but because the wanting-signal changed. Same input, different chemical mode, different engagement.
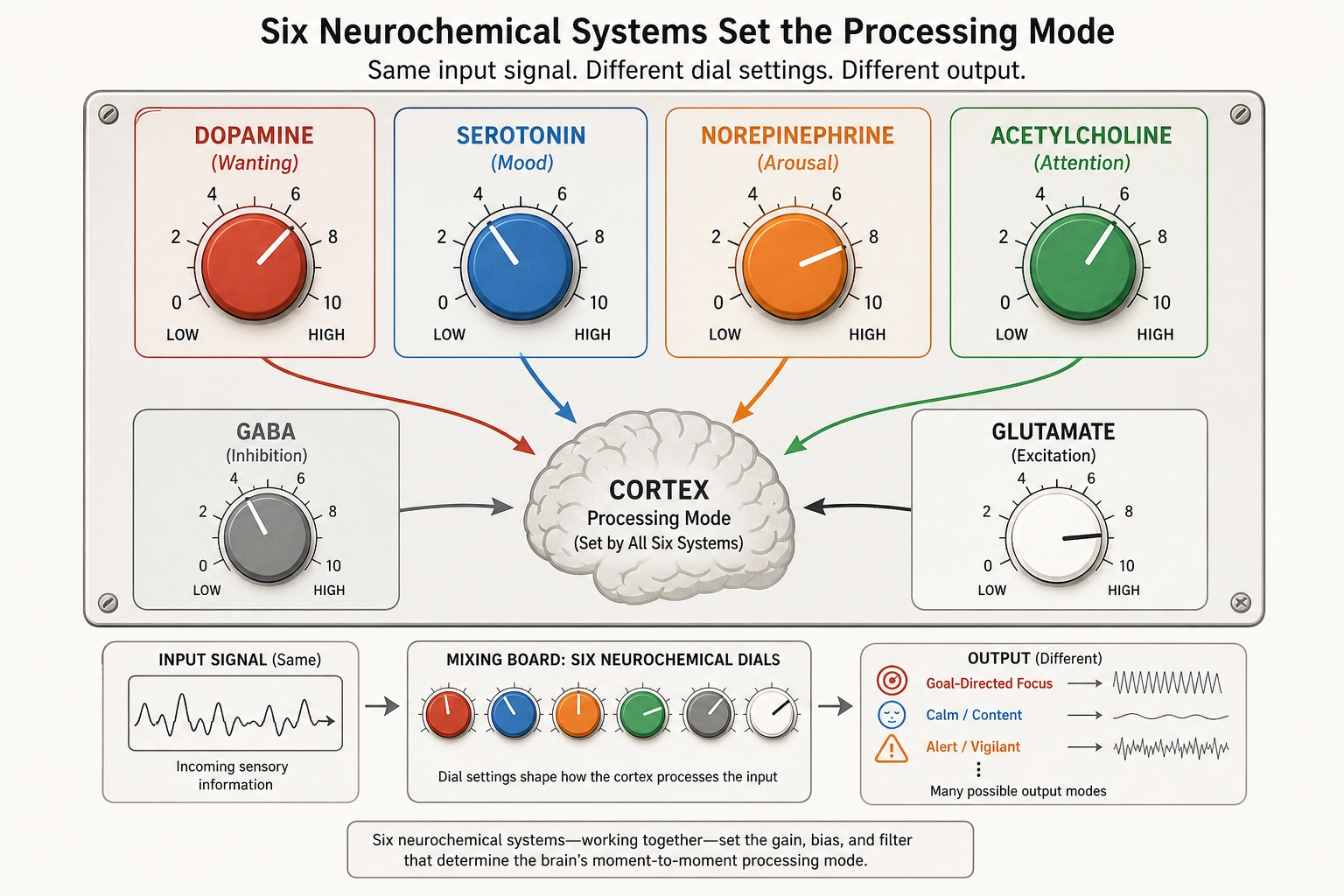
| System | Primary Source | Major Targets | Core Function | Classification (Tool 5) |
|---|---|---|---|---|
| Dopamine | VTA, substantia nigra (midbrain) | Prefrontal cortex, striatum, nucleus accumbens | Wanting, incentive salience, the motivational drive toward a stimulus, not pleasure itself | State producer (wanting) |
| Serotonin | Raphe nuclei (brainstem) | Widespread cortical and subcortical | Mood regulation, impulse control, emotional tone | Modulator |
| Norepinephrine | Locus coeruleus (pons) | Widespread cortical | Arousal and alertness, phasic bursts (brief, sudden) for salient events, tonic levels (sustained, ongoing) for exploration vs. exploitation | Modulator |
| Acetylcholine | Basal forebrain (nucleus basalis of Meynert) | Cortex, hippocampus | Attention, learning, memory encoding | Modulator |
| GABA | Distributed (~20% of cortical neurons) | Local circuits everywhere | Primary inhibitory neurotransmitter, suppresses neural activity by silencing competing signals | Infrastructure |
| Glutamate | Distributed (~80% of cortical neurons) | Local and long-range circuits everywhere | Primary excitatory neurotransmitter, drives neural activity | Infrastructure |
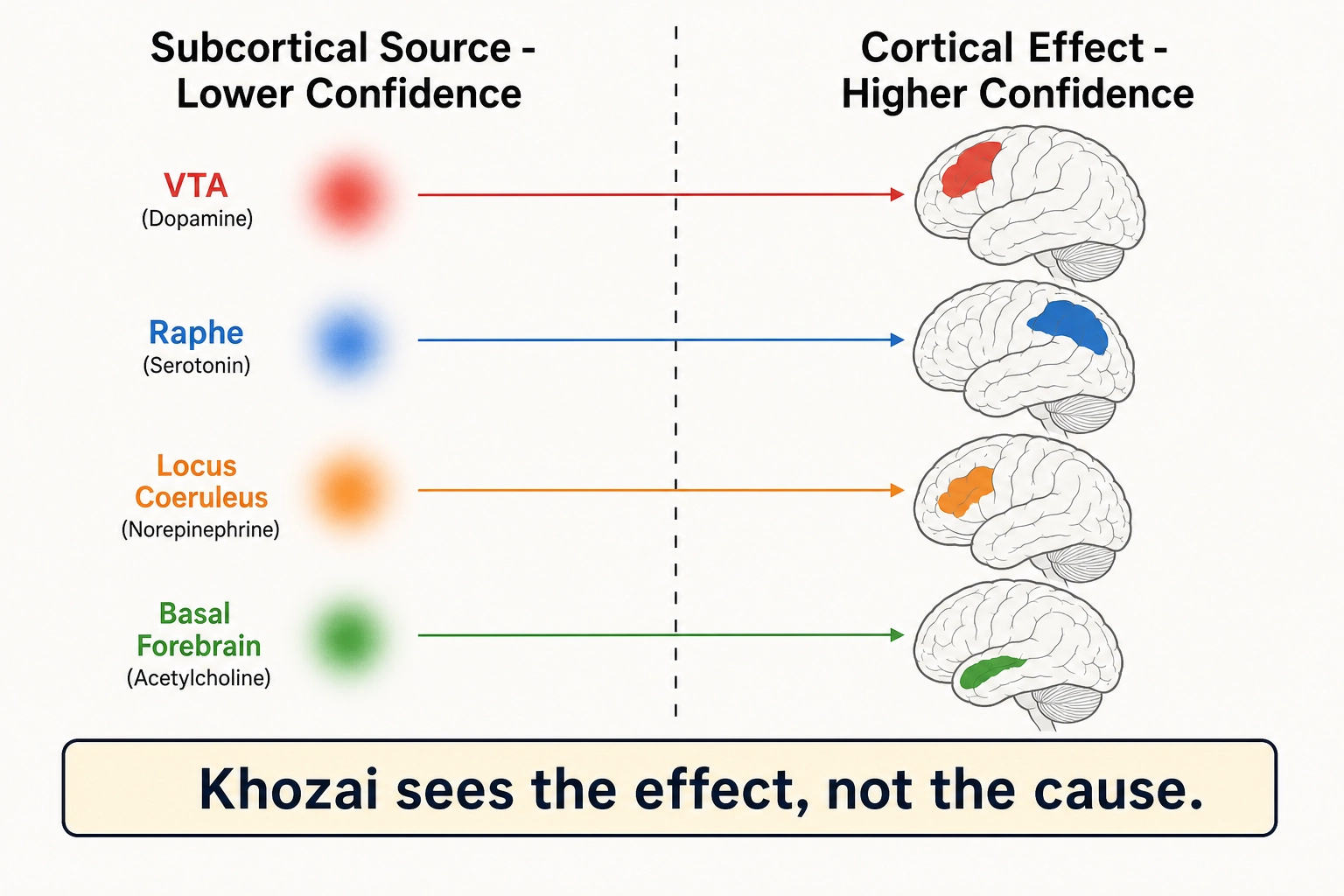
What Khozai can and cannot see in each system. Every chemical system listed above originates in small, deep brain structures, so the signal-quality gradient from Section 2.2 applies. But the effects of each system travel to the cortical surface, where predictions are more precise. The pattern is consistent: Khozai sees what the chemical systems do to the cortex, but cannot identify which chemical caused the change.
| System | Subcortical source (lower confidence) | Cortical effect (higher confidence) | What Khozai sees |
|---|---|---|---|
| Dopamine | VTA | Frontoparietal control network activation in motivated states | The cortical signature of wanting, not the dopamine release itself |
| Serotonin | Raphe nuclei | Shifted limbic and default mode network activation | The mood shift, not which chemical caused it |
| Norepinephrine | Locus coeruleus | Dorsal attention network (focused) vs ventral attention network (scanning) | The engagement mode, not the norepinephrine level |
| Acetylcholine | Basal forebrain | Changes in cortical response strength | The attention/learning effect, not the cholinergic mechanism |
| GABA / Glutamate | Distributed everywhere | Net activation pattern: which regions active, which quiet | The sculpted result, not the balance of excitation vs suppression |
Khozai sees the room temperature change; it cannot see which dial was turned.
Drs. Kent Berridge and Terry Robinson [5] (1998) demonstrated the double dissociation between wanting and liking in Brain Research Reviews. Dopamine depletion eliminates wanting, the motivational drive toward a reward, while leaving liking, the hedonic pleasure upon receiving it, intact. Liking depends on opioid systems, not dopamine. The wanting-liking dissociation has been replicated for over 25 years. For Khozai, this means the viewer’s motivational response (wanting to keep watching, wanting to share, wanting to act) and their hedonic response (enjoying what they see) run on separable neural systems - separable in the table above (dopamine → wanting via frontoparietal cortical signature) and separable in behavior.
Drs. Gary Aston-Jones and Jonathan Cohen [6] (2005) proposed the adaptive gain theory of the locus coeruleus-norepinephrine system in the Annual Review of Neuroscience. The locus coeruleus operates in two modes: phasic (brief bursts during salient events, promoting focused exploitation of current information) and tonic (sustained elevated firing, promoting broad exploration and distractibility). The cortical signatures - dorsal attention network in phasic/exploit mode, ventral attention network in tonic/explore mode - are the higher-confidence signals Khozai reads (see table above).
Chemistry modulates the brain’s processing mode. Three additional global properties constrain what that processing can produce: the brain’s alertness level, its use of suppression, and its hierarchical organization.
4. The Global Controls: Arousal, Inhibition, Hierarchy
4.1. Arousal: The Reticular Activating System
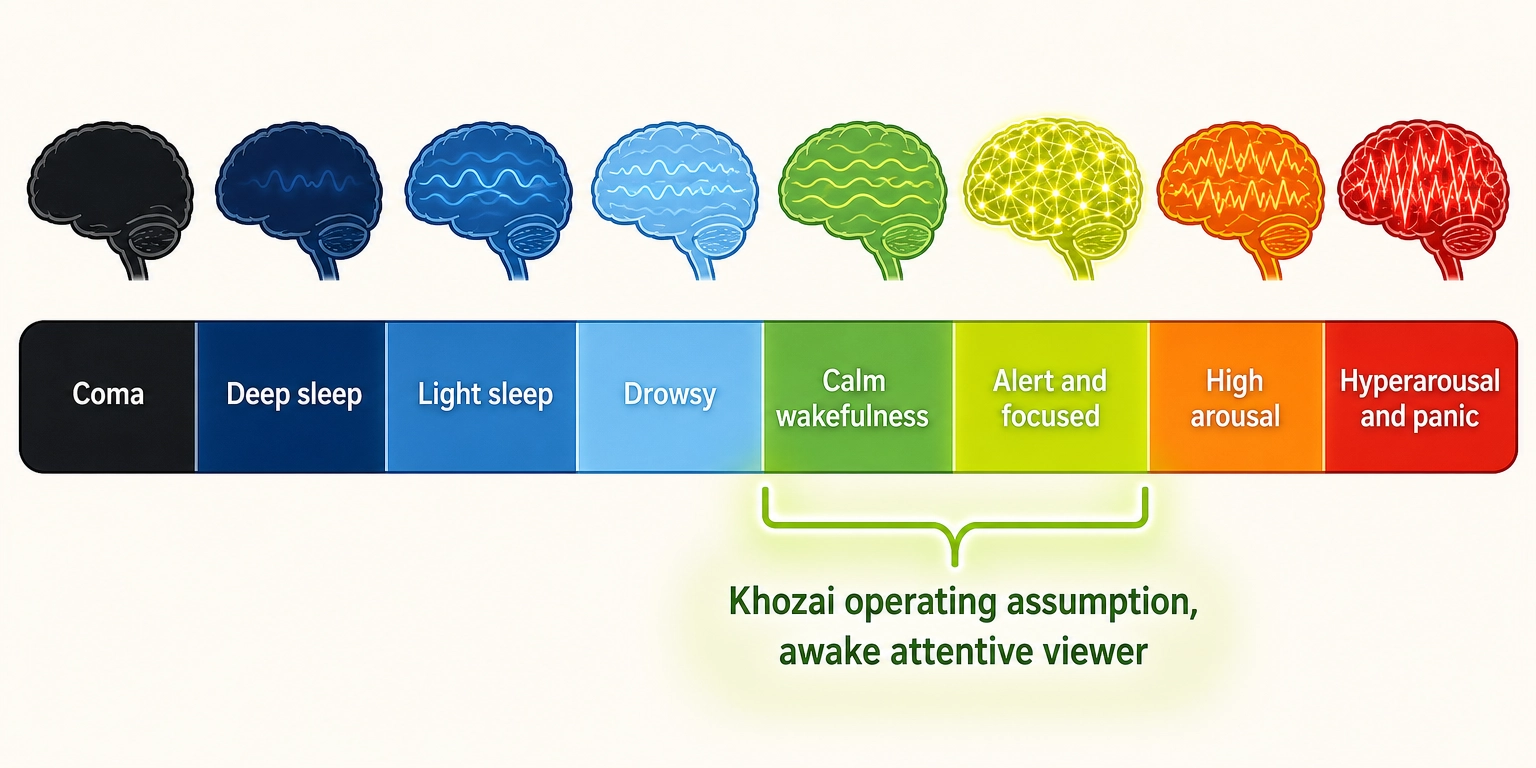
Property 5: The brain’s master alertness switch, the brainstem reticular activating system (RAS), controls the brain’s global level of activation.
The RAS is a spread-out network of neurons spanning the midbrain, pons, and medulla. It determines whether the brain is awake, drowsy, or comatose.
Strong evidence supports this. Drs. Giuseppe Moruzzi and Horace Magoun [7] (1949) showed in Electroencephalography and Clinical Neurophysiology that electrical stimulation of the brainstem reticular formation in anesthetized cats produced cortical waking patterns, while lesions produced permanent coma. In humans, severe RAS damage causes coma, the entire cortex goes dark regardless of its intrinsic wiring.
The RAS is a modulator. It does not produce any specific aspect of subjective experience. It sets the gain for all cortical processing. Without sufficient RAS activation, no content engagement is possible, not because the cortex is damaged, but because the cortex is not activated enough to process anything.
| Aspect | What Khozai sees | Limitation |
|---|---|---|
| RAS activation | Predicted at subcortical resolution (Section 2.2) | Cannot distinguish arousal levels finely |
| Cortical arousal effect | Widespread cortical activation vs deactivation | Sees the effect, not the brainstem mechanism |
| Viewer state | TRIBE v2 assumes an awake, attentive viewer - no input for drowsiness or distraction | If the viewer is drowsy, the prediction may not match the actual response |
| Mitigation | Khozai’s experimental pipeline (Chapter 8) can study arousal effects by varying viewer conditions | TRIBE v2’s limitation, not Khozai’s |
Dr. James Russell [8] (2003) proposed in Psychological Review that all emotional experience has two dimensions, each running from one extreme to the other, pleasure—displeasure and activation/arousal, forming what he called “core affect.” The arousal dimension of core affect is partly regulated by the RAS.
4.2. Inhibition
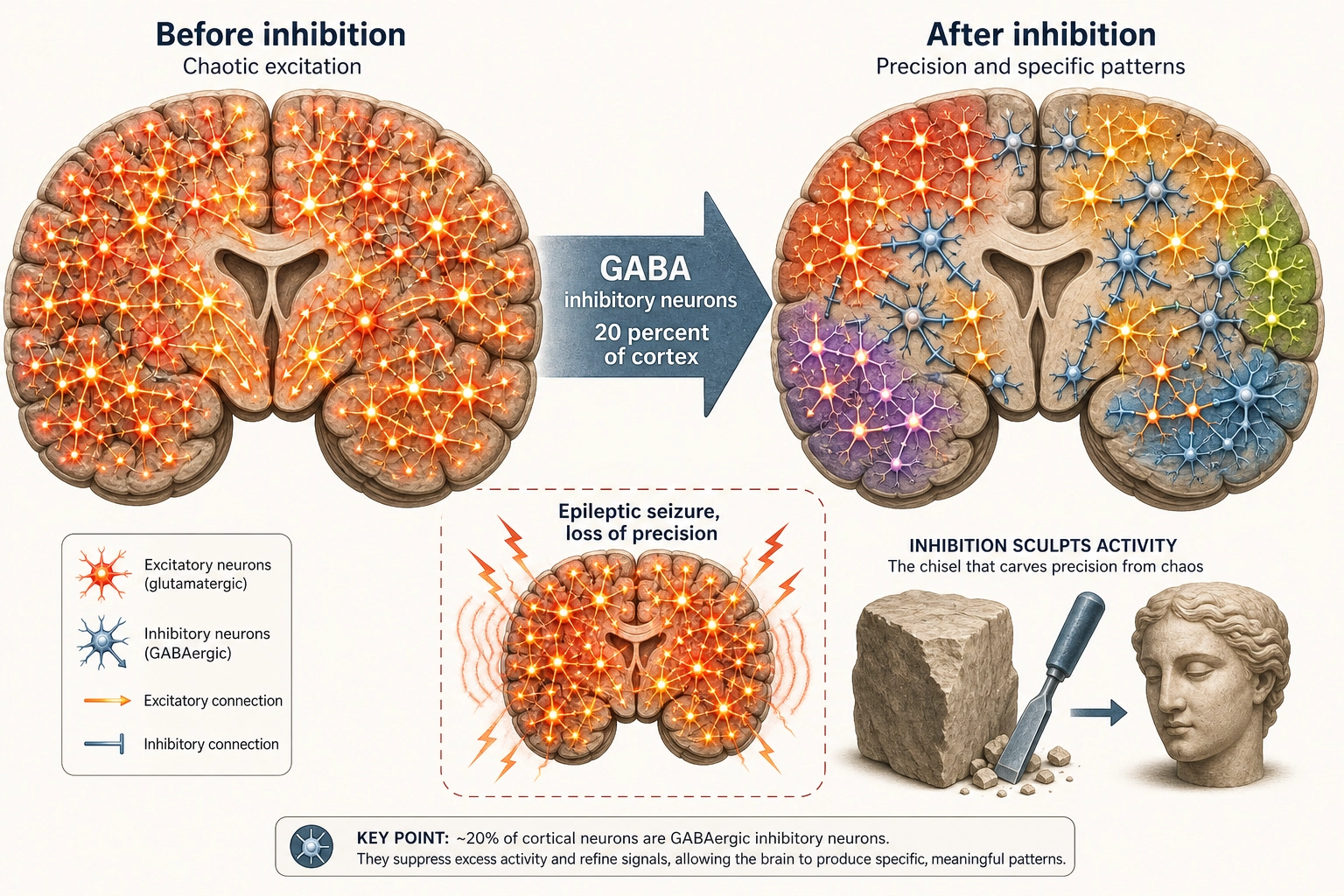
Property 6: Approximately 20% of cortical neurons are inhibitory. The brain shapes its responses by suppression, not just by activation.
The brain does not process information solely by turning things on. It processes by selectively turning things off. Inhibitory neurons, using GABA as their neurotransmitter, suppress the activity of nearby excitatory neurons. The result: out of all possible activations, inhibition carves a specific pattern by silencing everything except what is relevant.
What happens when inhibition fails? Epileptic seizures, chaotic, uncontrolled activation across entire brain regions. Not a loss of any specific experiential dimension, but a collapse of processing precision. This makes inhibition infrastructure under Tool 5: destroying it degrades everything without eliminating anything specific.
Khozai implication. Every Vₙ prediction (Khozai’s predicted whole-brain activation pattern for a piece of content) is a prediction about the net result, which regions ended up active and which ended up quiet, after excitation and inhibition have competed. Here is a concrete example: when a viewer watches a fast-action scene, TRIBE v2 might predict that V1 (primary visual cortex) is strongly active and the default mode network (self-referential thinking, mind-wandering) is deactivated. What happened inside the brain: excitatory neurons drove V1 hard because the visual input was intense, while inhibitory neurons suppressed the default mode network because the brain shifted resources away from daydreaming toward processing the action. Khozai sees the result of this competition, V1 up, default mode down, but cannot tell how much of each pattern was driven by activation versus shaped by suppression. It sees the sculpted statue, not the chisel strokes.
4.3. Hierarchical Processing
Property 7: Cortical processing is organized hierarchically, from simple features to complex representations, with both feedforward and feedback connections.
The visual system is the best-studied example. Raw visual signals from the retina arrive at V1, where neurons respond to simple features: edges at specific orientations, small patches of contrast. From V1, information flows forward through a series of areas. Each level builds more complex representations from the simpler ones below.
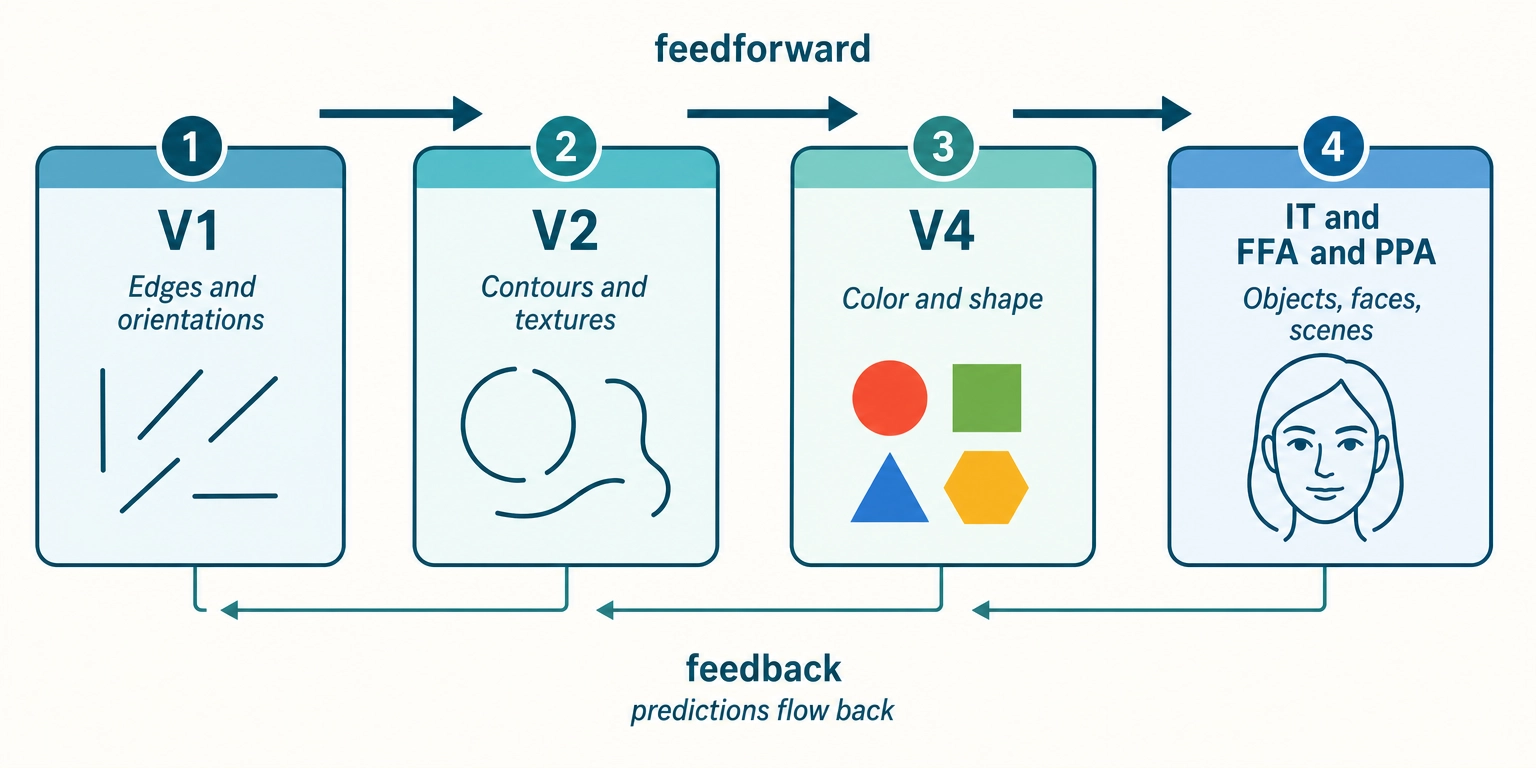
The feedforward direction builds representations: edges become contours, contours become shapes, shapes become objects and faces. The feedback direction carries predictions and contextual information: higher areas tell lower areas what to expect, and lower areas send back prediction errors, the mismatches between what was expected and what was received.
Dr. Karl Friston [9] (2010) formalized this as the predictive processing framework in Nature Reviews Neuroscience. The core idea: higher cortical levels generate predictions about incoming sensory data. Lower levels compare those predictions to actual input and propagate only the difference, the prediction error, upward. The brain is not passively receiving input. It is actively predicting and updating only when surprised. The predictive processing framework, while influential, remains debated: critics have raised the ‘dark room problem’ (why organisms do not simply seek stimulus-free environments) and questioned whether the framework generates falsifiable predictions at the neural level (Anderson and Chemero, 2013). The framework treats predictive processing as a useful organizing principle for understanding content engagement, not as a settled neuroscientific consensus.
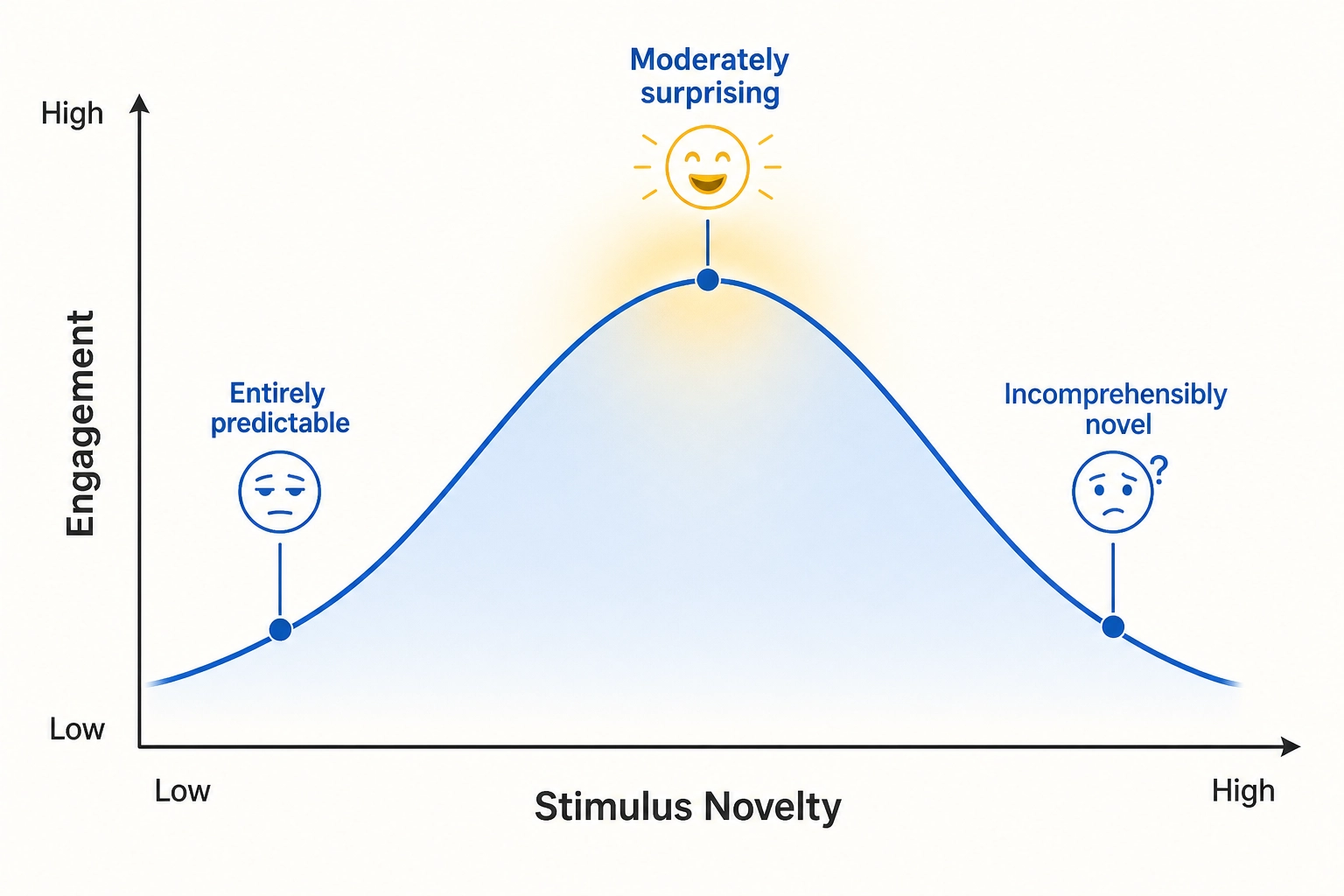
For Khozai, this changes what a cortical response means. A viewer’s response to content is not just “what the content contains.” It is “how the content differs from what the brain expected.” Content that is entirely predictable produces minimal prediction error and minimal cortical response. Content that is moderately surprising produces strong engagement. Content that is incomprehensibly novel produces confusion and disengagement.
Dr. Daniel Berlyne [10] (1960) described exactly this pattern as the inverted-U relationship between stimulus novelty and interest in Conflict, Arousal, and Curiosity. Moderate novelty is optimally engaging. Khozai’s framework connects this behavioral observation to a neural mechanism: the prediction-error hierarchy in cortical processing. Recent empirical work has confirmed this link with both behavioral and neural evidence. The neuroscientist Benjamin Gold and colleagues [16] (2019) showed in the Journal of Neuroscience that musical sequences with intermediate predictability produced the strongest pleasure ratings, forming an inverted-U curve over prediction error magnitude. The neuroscientist Vincent Cheung and colleagues [17] (2019) demonstrated in Current Biology that musical pleasure peaks when uncertainty and surprise interact at intermediate levels, with corresponding activation in the amygdala, hippocampus, and auditory cortex. Together, these studies provide direct evidence that moderate prediction error produces the strongest engagement, bridging Berlyne’s behavioral observation to the predictive processing framework.
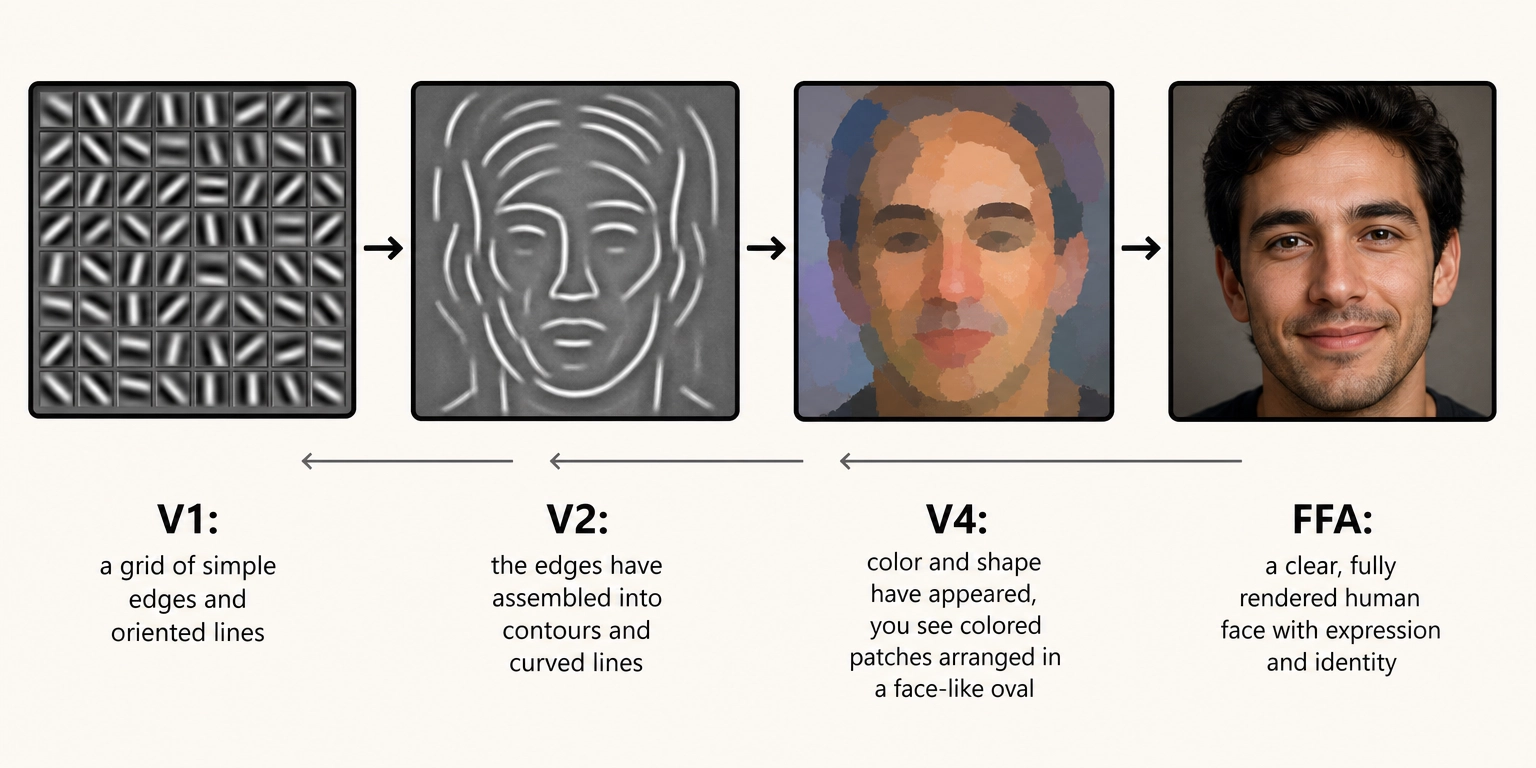
Hierarchical processing is infrastructure under Tool 5. Disrupting it degrades processing quality at a specific level, it does not eliminate a specific aspect of subjective experience. A lesion to V4 eliminates color perception specifically (achromatopsia), but that is a loss of V4’s state-producing function as a region, not a loss of hierarchy as a principle. The hierarchy is the organizational scheme that makes it possible for specific regions to have specific functions.
Khozai implication. TRIBE v2 predicts activation at every level of the hierarchy simultaneously. This makes the activation pattern directly interpretable. Khozai can observe whether a piece of content engages early visual processing (high V1/V2 activation, rich visual detail or texture), late visual processing (high FFA/PPA activation, face or scene recognition), or both. Activation at different hierarchy levels means different things about the viewer’s experience. For example: if TRIBE v2 predicts high V1/V2 activation but low FFA activation, the content is visually rich (complex textures, strong contrasts) but does not engage face processing, think of a nature documentary landscape shot. If it predicts low V1/V2 but high FFA, the content is visually simple but face-driven, think of a close-up interview on a plain background. Each pattern tells Khozai something different about what the viewer is processing.
The properties described so far - receptors, regions, networks, chemistry, and global controls - are the brain’s processing machinery. Two additional properties connect that machinery to the body and to its own history.
5. The Body Connection and Plasticity
5.1. The Body Connection
Property 8: The brain is connected to the body through its body-monitoring center (the hypothalamus) and internal-organ sensors (visceral afferents). It continuously monitors the body’s internal state.
The brain does not float in isolation. The hypothalamus, a small subcortical structure, monitors and regulates body temperature, hunger, thirst, hormonal balance, and circadian rhythm (the body’s internal 24-hour clock). Visceral afferents (receptor system #10 from the table in Section 1) report the state of internal organs back to the brain through the brainstem and thalamus, reaching cortical representation in the insular cortex.
Khozai implication first: body-state monitoring is primarily subcortical (hypothalamus, brainstem), so the same signal-quality gradient from Section 2.2 applies. The cortical representation of body state in the insular cortex provides a complementary, higher-resolution signal.
| Stage | Where | Khozai’s confidence | What Khozai sees |
|---|---|---|---|
| Body organs → brainstem | Subcortical | Lower | Blurred - how raw signals were filtered and prioritized |
| Brainstem → thalamus → posterior insula | Mixed | Lower → Higher | Transition zone |
| Anterior insula (cortical) | Cortical surface | Higher | Clear: final integrated representation of body state |
Dr. A.D. (Bud) Craig [11] (2002) described in Nature Reviews Neuroscience how this pathway works. Interoceptive signals, the sense of the body’s physiological condition, ascend from the body through the brainstem and thalamus to the posterior (rear) insula, then to the anterior (front) insula, where they are integrated with emotional and cognitive context.
Dr. António Damásio [12] (1994) proposed the somatic marker hypothesis in Descartes’ Error. The claim: body-state signals, felt as gut feelings, background moods, and visceral responses, bias decision-making and shape emotional experience. The body is not a passive vehicle for the brain. It is an active contributor to what the viewer feels.
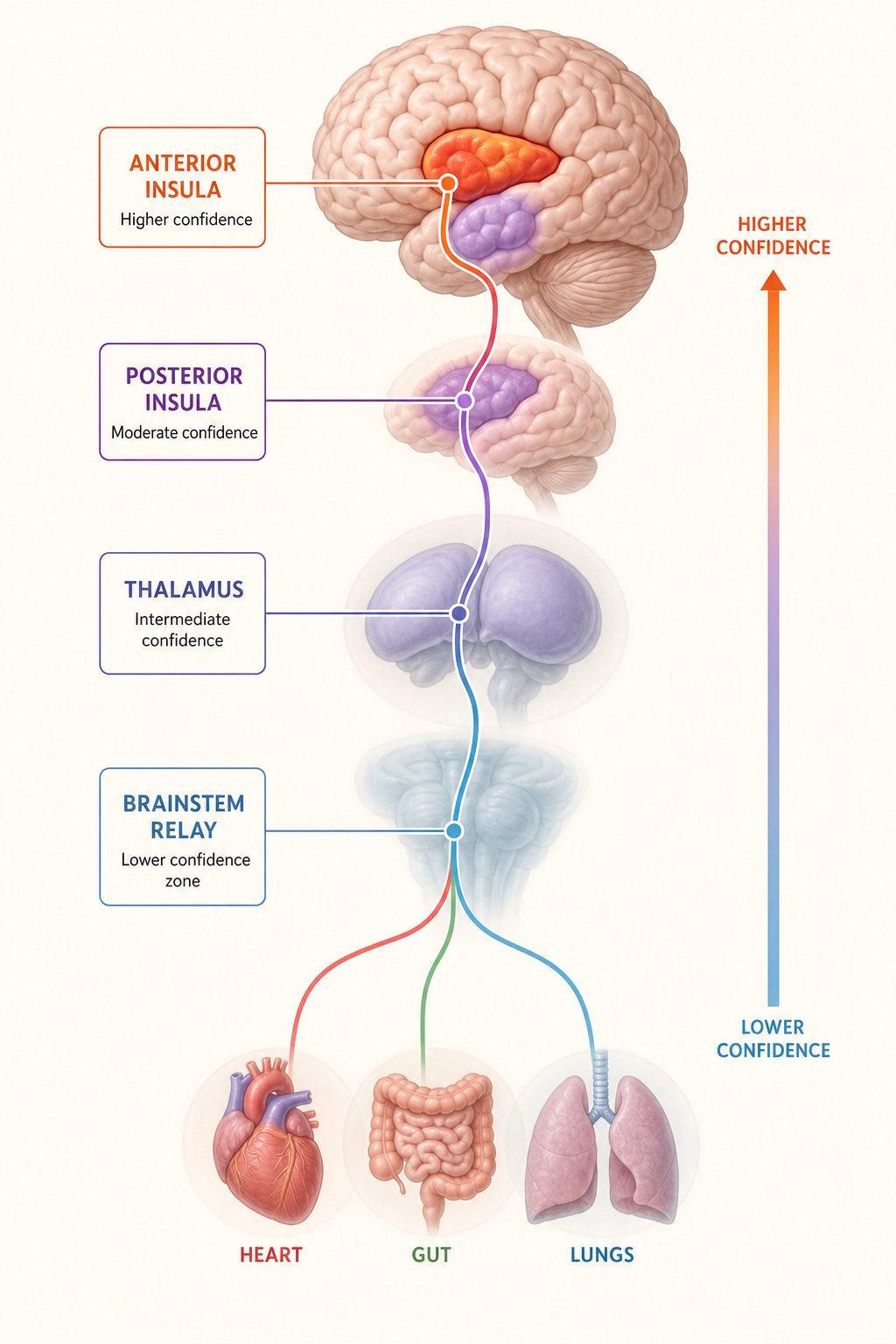
What can Khozai see here? When TRIBE v2 predicts strong insular cortex activation, Khozai can infer that the content likely engages interoceptive or emotional-awareness processing. What it cannot determine is whether this reflects actual body-state changes or purely top-down cortical activation.
The body connection is a state producer at the experiential level. Spinal cord transection eliminates bodily awareness (the “Bodily” dimension in Experience Space). Insular cortex lesions reduce the sense of one’s own physiological condition. Destroying the body-monitoring system removes a specific aspect of subjective experience.
5.2. Plasticity
Property 9: The brain reshapes itself with use. Connections that fire together strengthen; connections that do not fire together weaken.
This is Hebbian plasticity, named after the psychologist Donald Hebb’s [13] 1949 principle: “neurons that fire together wire together.” Synapses, the connection points between neurons, strengthen when the sending neuron and the receiving neuron are active at the same time, and weaken when they are not. This is not a metaphor. It is a measurable biological process, long-term potentiation and long-term depression.
The consequence: the brain is not a fixed circuit. Every time the viewer watches content, their brain changes slightly. Repeated exposure strengthens the connections between neurons involved in processing that content, the synapses get more efficient, and the chain of regions that processed the content becomes faster and more responsive on the next encounter. This is how preferences form, how expertise develops, and how habituation occurs.
In Khozai’s formal framework, plasticity is Mapping 5, the feedback loop from Experience Space back to Neural State Space. Attending to a stimulus changes how that stimulus is processed on the next encounter. The viewer is not a fixed receiver. They are a system that reshapes itself in response to what it processes.
Plasticity is infrastructure under Tool 5. It does not produce any specific aspect of subjective experience, it changes the efficiency and pattern of processing over time. Removing plasticity completely would not eliminate any experiential dimension. It would prevent the brain from learning, adapting, or forming memories.
Khozai implication. TRIBE v2 predicts the response of an average subject at a single point in time. The encoding model has no input for “how many times has this viewer seen this content before”, so its prediction is always a first-exposure prediction.
But Khozai the system is not limited to first-exposure analysis. Khozai’s mutation and measurement pipeline (Chapters 7 and 8) can study plasticity directly: show the same content to viewers who have seen it 0, 1, 5, or 10 times, measure their behavioral responses (Vₚ, the platform metrics like views, retention, likes, and shares described in Chapter 2) for each group, and correlate exposure count with engagement changes. Repetition count becomes a variable in the experimental design, just like any other variable. What TRIBE v2 cannot model, Khozai’s correlation engine can discover empirically.
Properties 1 through 9 describe structures and systems - the hardware itself. Four additional properties describe how that hardware is organized.
6. The Architectural Principles
Properties 10 through 13 are not structures or systems. They are architectural facts about how the brain is organized. They constrain what any measurement or prediction system can do, including Khozai.
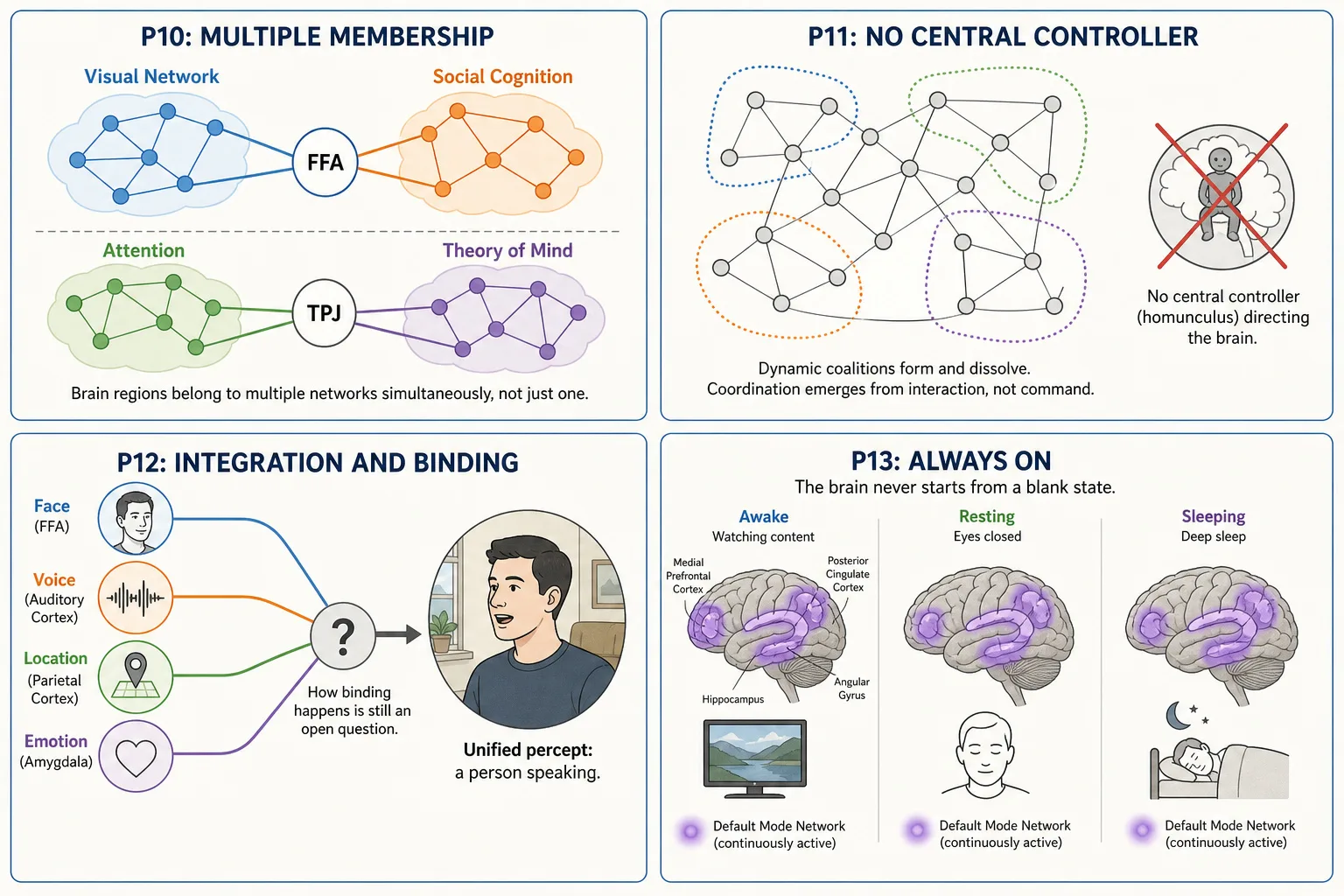
| Property | Principle | What It Means | What Breaks If Ignored |
|---|---|---|---|
| P10: Multiple Membership | Every cortical region belongs to multiple functional networks simultaneously | The FFA (fusiform face area) is part of the visual network AND participates in social-cognition processing. The TPJ (temporal parietal junction) contributes to both the ventral attention network and theory-of-mind processing. Regions are not exclusively assigned to one function. | Interpreting Vₙ as if each region had one function would produce systematically wrong inferences. A region’s activation must be interpreted in the context of what else is co-activated. |
| P11: No Central Controller | There is no single brain region that “decides” or “controls” everything | No homunculus (no little person inside the brain making the decisions). No executive region that all information flows through. Coordination emerges from network interactions, dynamic coalitions of regions that temporarily synchronize for a task and then reconfigure. | Assuming a central decision point would lead to looking for one region that “explains” engagement. No such region exists. Engagement is a distributed pattern across multiple networks. |
| P12: Integration / Binding | The brain produces unified subjective experience from distributed processing across many separate regions | The viewer sees a face with a voice at a location, a unified percept. The underlying processing is distributed: face processing in FFA, voice processing in auditory cortex, spatial processing in parietal cortex, emotional evaluation in amygdala. How these are bound into one experience is the binding problem, one of the open questions in neuroscience. | Treating each Vₙ region prediction as an independent channel would miss the fact that the viewer’s experience is unified. A face + a voice + an emotion is not three separate experiences; it is one integrated experience with three identifiable components. |
| P13: Always-On | The brain never fully turns off, even during sleep | The default mode network is active during rest. Sleep has multiple stages with distinct neural processing. Even under anesthesia, some neural activity continues. The brain is always processing, predicting, and maintaining. | The viewer brings their current mental state to every piece of content. TRIBE v2 has no input for that state, it takes only the video file. Its prediction is most accurate for viewers in a typical, unremarkable state, and less accurate for viewers in unusual states. |
P13 extended note. The always-on property also means the viewer’s actions, scrolling, pausing, rewatching, reshape their stimulus environment, producing new inputs for the brain (Mapping 5: Behavioral Output to Physical Stimulus Space feedback). Assuming the brain starts from a “blank state” when content begins would ignore the viewer’s ongoing cognitive and emotional processing. TRIBE v2’s prediction is the same whether the viewer is alert or drowsy, happy or anxious, just ate or is hungry.
These four architectural principles (Properties 10—13) together describe a brain that is distributed, always-active, multiply-connected, with no central controller and unified experiential output.
This is not a complication to work around. It is the architecture that makes Khozai’s approach possible.
Because the brain is distributed and multiply connected, a change in one content property (say, adding a face to a scene) activates a specific set of regions (FFA, TPJ, amygdala) without disrupting the rest. If the brain were a single undifferentiated processor, changing one content property would change the whole response and nothing could be traced. Because there is no central controller, the activation patterns from different content properties do not all funnel through one bottleneck region where they would merge and become inseparable. Each pattern retains its identity. Because the brain is always on, Khozai knows the viewer is never a blank receiver, they bring ongoing thoughts, moods, and expectations that shape how the content is processed, which is why TRIBE v2’s prediction (which has no input for the viewer’s state) is an approximation, not a perfect readout.
These thirteen properties - nine hardware, four architectural - are the ones Khozai’s predictions depend on. Eight additional properties affect the brain’s operation but are not catastrophic for the framework if omitted.
7. Secondary Properties
The thirteen core properties above are the ones whose removal would break Khozai’s predictions in a specific, traceable way. The eight secondary properties below are real features of the brain that affect its operation but are not catastrophic for Khozai if omitted from the model.
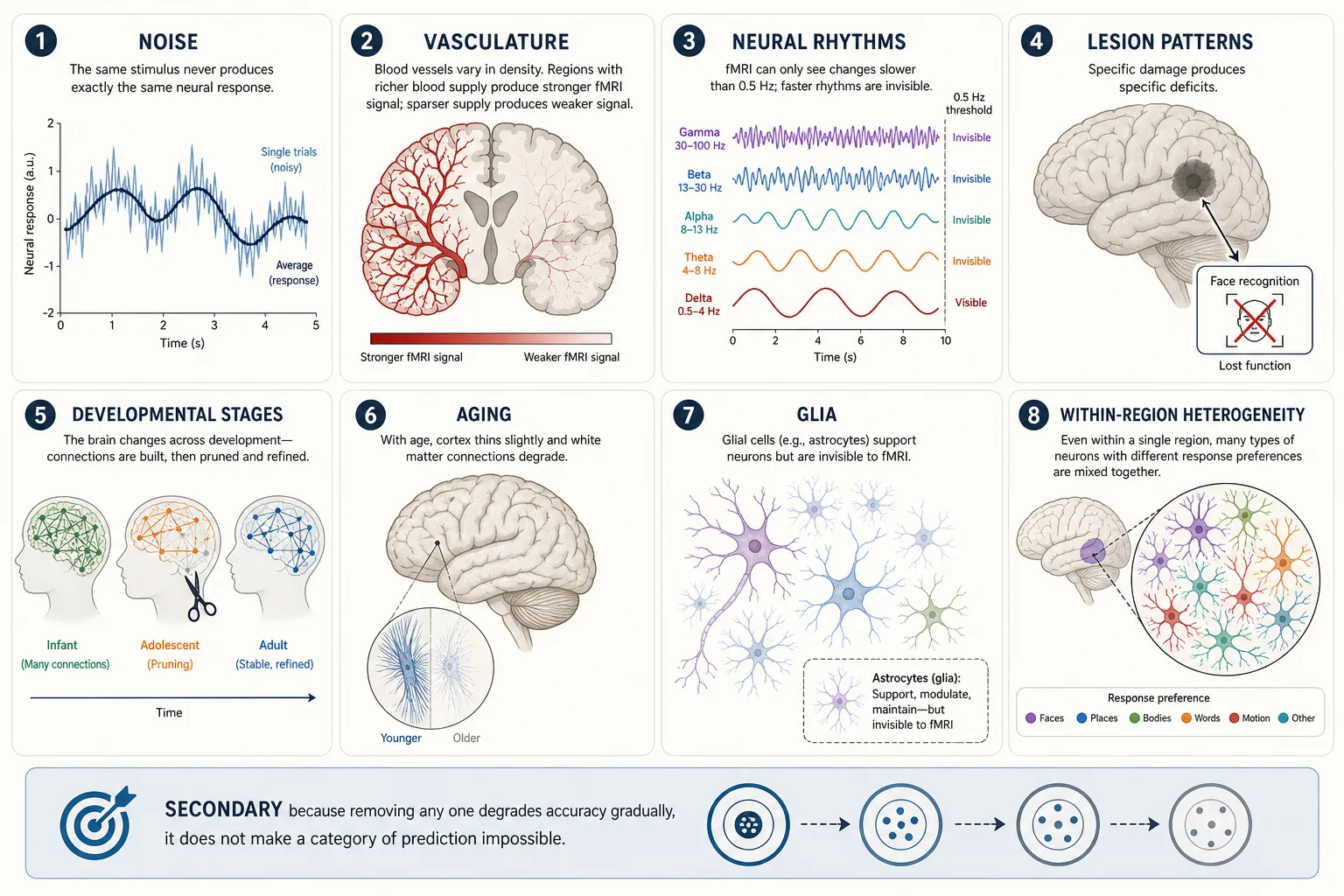
| # | Property | What It Is | Khozai Implication |
|---|---|---|---|
| 1 | Noise | Neural firing is inherently stochastic (random from one moment to the next), the same stimulus never produces exactly the same neural response twice. Trial-to-trial variability is a fundamental property of neurons, not measurement error. | TRIBE v2 predicts the average response across subjects. The noise in individual responses is averaged out. This makes TRIBE v2’s prediction more stable but also means it does not capture individual variation. Khozai’s experimental pipeline can study individual differences through behavioral data (Vₚ), even though TRIBE v2 cannot. |
| 2 | Vasculature | The brain’s blood supply determines which regions receive oxygen and glucose. fMRI does not measure neural activity directly, it measures blood-oxygenation changes (the BOLD signal). When neurons fire, nearby blood vessels deliver extra oxygenated blood about 1—2 seconds later. fMRI detects that delayed blood-flow surge, not the neural firing itself. | TRIBE v2’s predictions are predictions about these blood-flow changes, not about the neural firing that caused them. This matters because different brain regions have different blood-supply densities, a region with richer blood supply produces a stronger BOLD signal even if its neural activity is the same as a less vascularized region. So some of what looks like “more activation” in Khozai’s predictions is actually “better blood supply.” This is a known limitation of all fMRI-based methods, not specific to Khozai. |
| 3 | Neural rhythms / oscillations | Neurons do not fire at a steady rate, they pulse in rhythmic waves, like an orchestra where different sections play at different tempos simultaneously. These rhythms are measured in Hz (cycles per second). Slow rhythms: delta (1—4 Hz, deep sleep), theta (4—8 Hz, memory encoding, e.g., the hippocampus pulses at theta when forming a new memory). Fast rhythms: alpha (8—13 Hz, relaxed wakefulness, close your eyes and alpha waves increase), beta (13—30 Hz, active thinking), gamma (30—100 Hz, focused attention, when you concentrate on a specific detail, gamma activity spikes in the relevant cortical region). These rhythms matter because they reflect moment-to-moment shifts in what the brain is doing: a shift from alpha to gamma in visual cortex means the viewer just went from passively watching to actively scrutinizing something (a transition documented with intracranial recordings by the neuroscientist Pascal Fries [18], 2015, Neuron). | TRIBE v2 was trained on fMRI data, and fMRI’s blood-flow changes lag 1—2 seconds behind neural activity. Oscillations faster than ~0.5 Hz were never in the training data. Khozai’s Vₙ predictions therefore cannot capture these rapid rhythmic shifts, they show where activity is, but not how it pulses moment to moment. |
| 4 | Lesion patterns | Specific lesions produce specific deficits (Premise 5). The pattern of deficits maps the structure-experience relationship. Akinetopsia (loss of motion perception from MT/V5 lesion; the neuropsychologist Josef Zihl, the neurologist Detlev von Cramon, and the anatomist Norbert Mai [14] 1983), prosopagnosia (loss of face recognition from FFA lesion), Broca’s aphasia (loss of speech production from inferior frontal lesion). | Lesion evidence is the empirical foundation for Experience Space dimensions (Chapter 4). It validates the interpretation of Vₙ predictions: if FFA lesion eliminates face recognition, then FFA activation predicts face processing. |
| 5 | Developmental stages | The brain develops through critical periods with different properties at each stage. Infant cortex is hyperplastic; adolescent cortex undergoes major pruning; adult cortex is relatively stable. | TRIBE v2 was trained on adult fMRI data. Its predictions may not generalize to children’s or adolescents’ brains. Khozai’s framework assumes adult viewers. |
| 6 | Aging | Cortical thinning, white-matter degradation, and neurochemical decline accumulate with age. Processing speed decreases. Some compensatory reorganization occurs. | TRIBE v2’s average-subject prediction does not model age-related variation. Khozai’s predictions may be less accurate for elderly viewers. |
| 7 | Glia | Glial cells (astrocytes, oligodendrocytes, microglia) outnumber neurons and perform support functions: metabolic supply, myelination, immune defense, synaptic modulation. | Glia affect neural processing but are invisible to fMRI. TRIBE v2’s predictions implicitly include glial effects (they’re part of the training data) but cannot separate them. |
| 8 | Within-region heterogeneity | Even within a single Glasser region or Yeo network, neurons have diverse response properties. The FFA contains neurons that respond to faces, but also neurons that respond to other complex visual stimuli. | Khozai operates at region and network level, not neuron level. Within-region heterogeneity is averaged out. This is acceptable at Khozai’s operational resolution but means the framework cannot distinguish between two stimuli that activate the same region with different within-region patterns. |
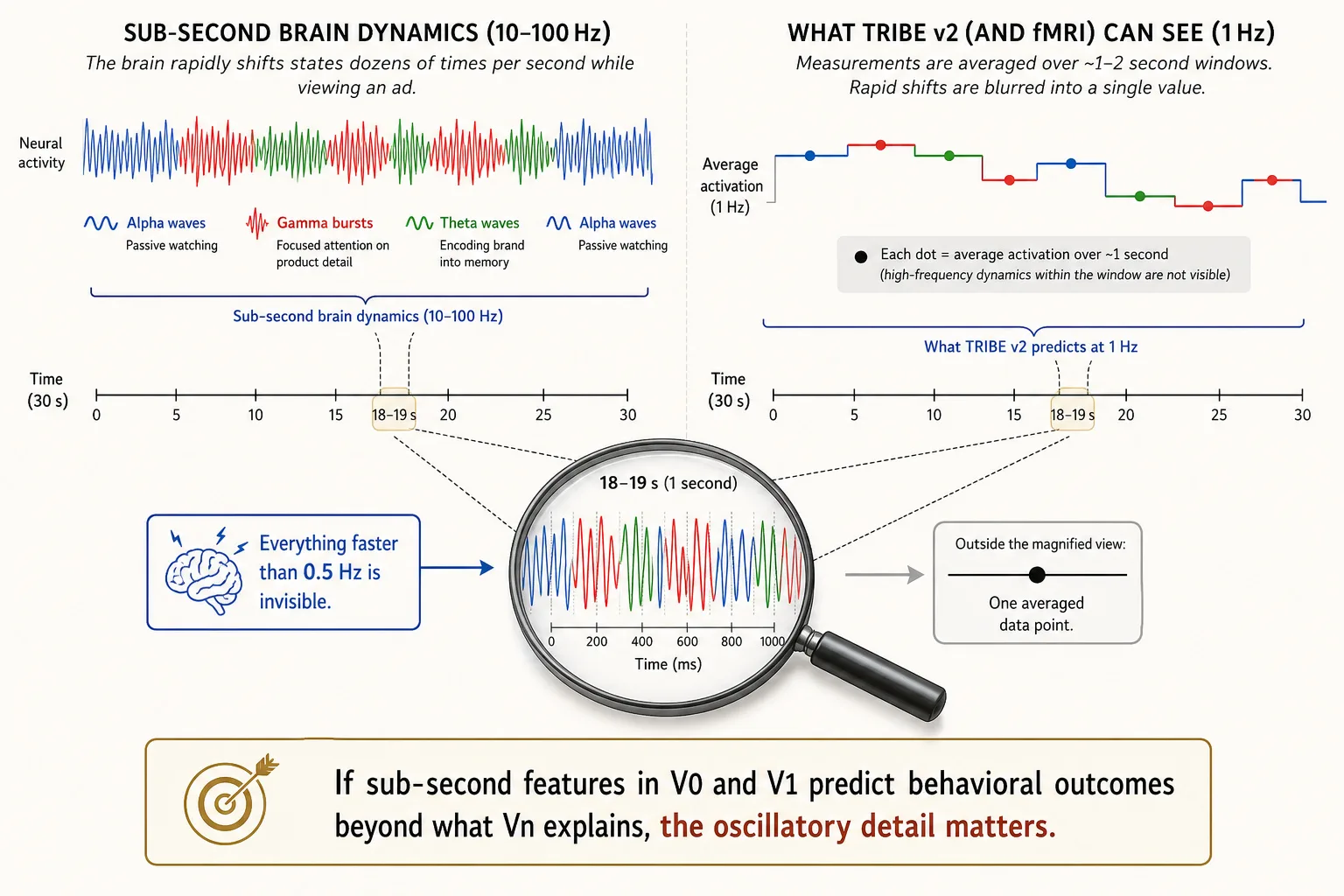
Neural rhythms, extended Khozai implication. Why does this matter for content? Consider a 30-second ad. In those 30 seconds, the viewer’s brain may shift between alpha (passively watching), gamma (focusing on a product detail), theta (encoding the brand into memory), and back, dozens of times. These shifts happen at the scale of hundreds of milliseconds. fMRI, which TRIBE v2 learned from, can only detect changes that last at least 1—2 seconds. Everything faster is invisible.
Khozai does not use a scanner, it predicts from the content file alone. TRIBE v2 outputs one prediction per second (1 Hz), so Khozai CAN tell you that at second 12 the viewer’s visual cortex spiked, at second 15 it dropped, and at second 18 it recovered. That second-by-second tracking is enough to map engagement across a video. What Khozai CANNOT capture is what happens WITHIN each second, the rapid oscillatory shifts (alpha to gamma to theta at 10—100 cycles per second) that reflect moment-to-moment changes in processing mode. Those sub-second dynamics are blurred away because fMRI, which TRIBE v2 learned from, averages over ~2-second windows.
If oscillatory information turns out to matter for content engagement, a complementary approach would be needed. As of May 2026, no encoding model exists that predicts EEG responses (millisecond-scale electrical brain activity measured from sensors on the scalp) from a content file the way TRIBE v2 predicts fMRI responses. EEG signals are noisier and harder to align across subjects than fMRI. Building an EEG-based equivalent of TRIBE v2 is an open research problem, not an available tool.
These eight secondary properties are not ignored. They constrain how confident Khozai can be and where predictions are likely to break down. They are secondary because removing any one from the model degrades accuracy gradually, it does not make a category of prediction impossible.
8. What the Architecture Means for Khozai
The brain described in this chapter has three properties that make Khozai’s approach viable.
Finite. Receptor systems: 10. Cortical regions: ~360. Neurochemical systems: 6 major. Functional networks: 7 or 17. Subcortical structures: finite. Finite means catalogueable. Catalogueable means measurable. This grounds Premise 2 (receptors), Premise 3 (cortical organization), and Premise 4 (subcortical structures).
Hierarchically organized. Cortical processing is arranged in levels, from simple features (V1) to complex representations (FFA, PPA, TPJ). Functional networks group regions by correlated activity. Experiential dimensions nest: vision contains color, motion, face recognition, and scene recognition as sub-dimensions. Hierarchical means decomposable. Khozai can analyze content effects at multiple resolutions (Tool 7) and trace specific behavioral outcomes to specific levels of the processing hierarchy.
Specific. Alter a specific structure, and a specific aspect of subjective experience changes while others remain intact (Premise 5). This specificity, the structure-experience mapping, is the foundation for everything that follows. Without it, the brain would be an undifferentiated mass where nothing could be traced, predicted, or isolated. With it, Khozai can predict that a specific content property will activate a specific cortical region, which contributes to a specific aspect of the viewer’s experience, which correlates with a specific behavioral outcome.
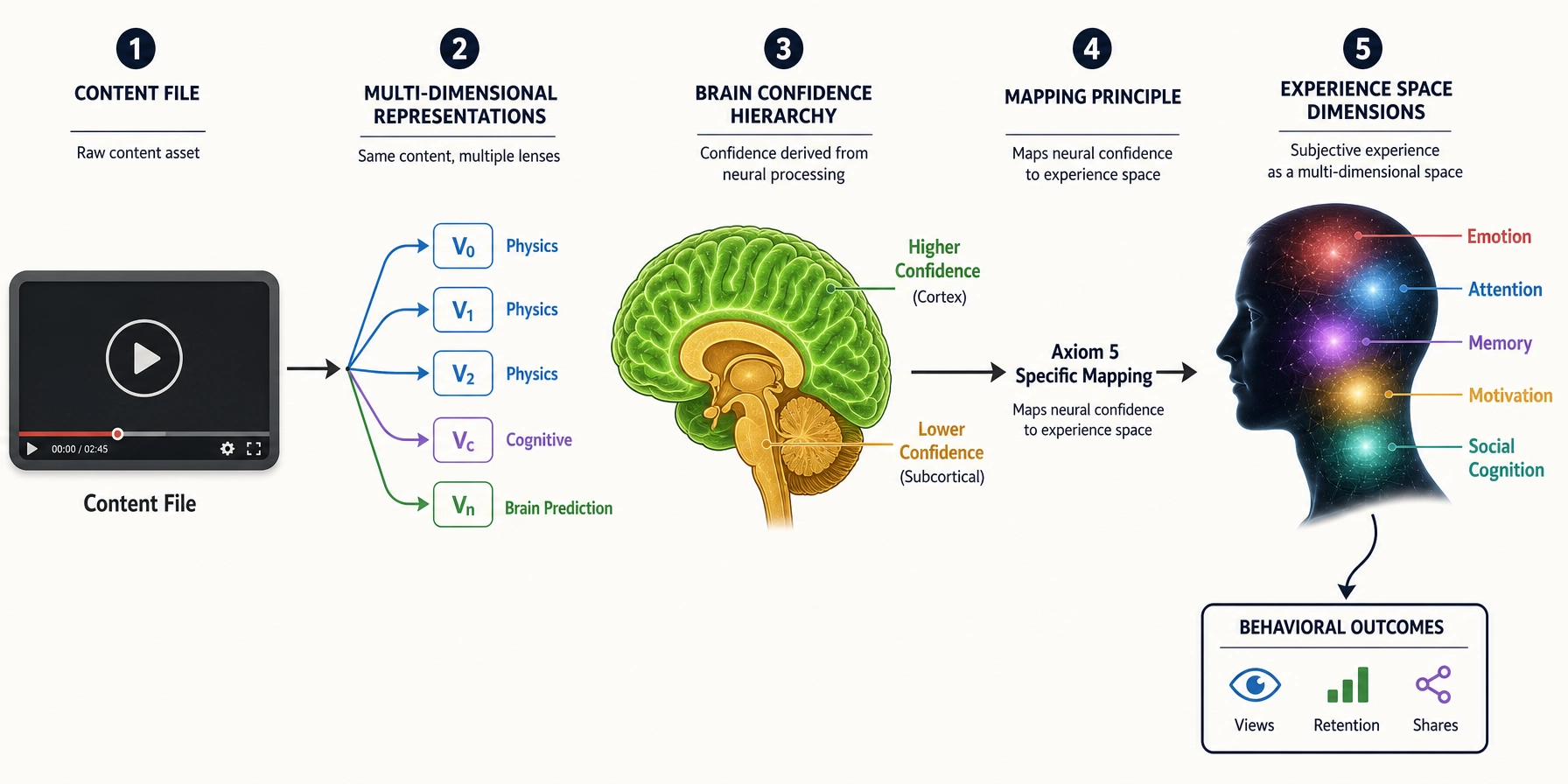
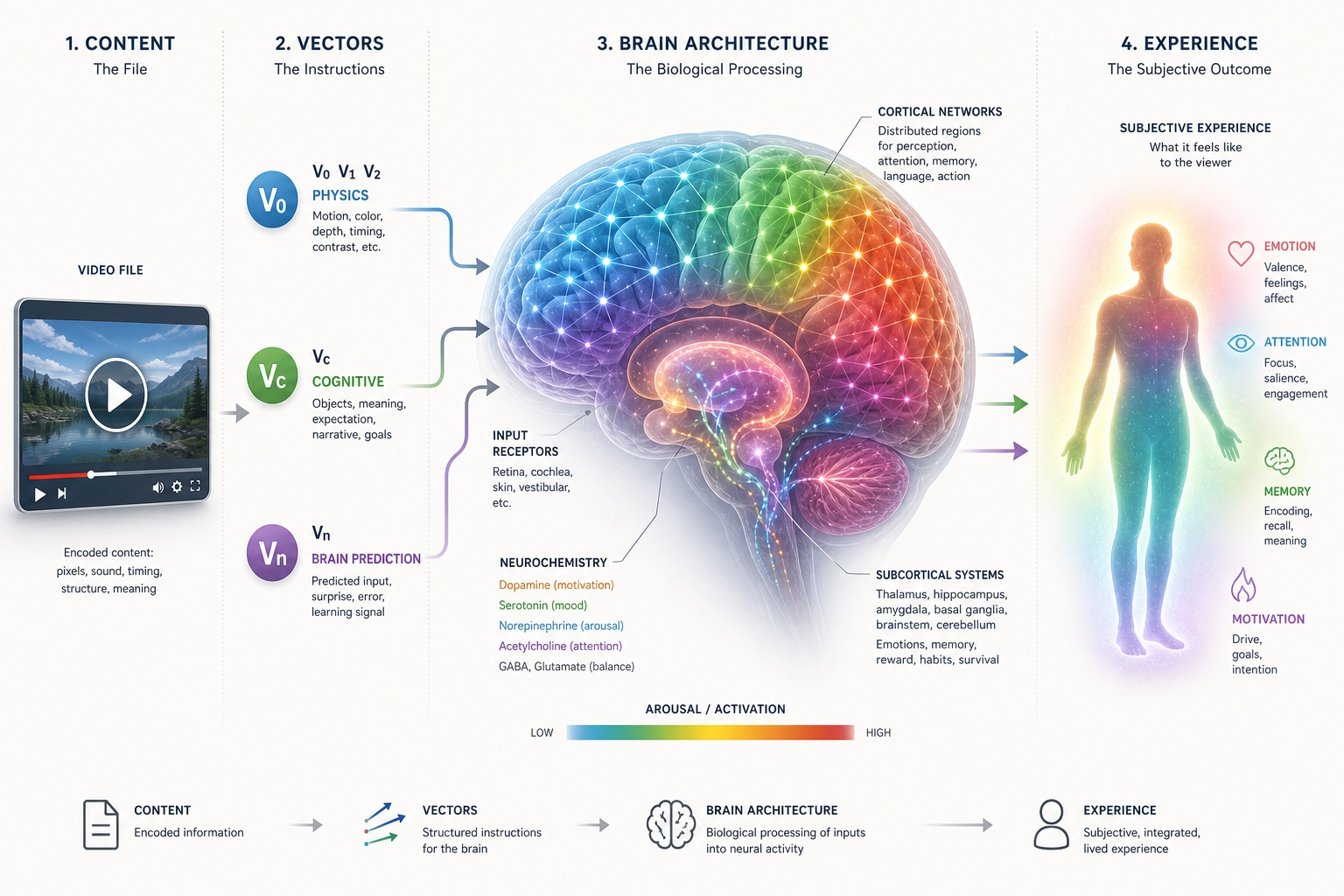
The chain from content to brain to subjective experience to behavior depends on every property in this chapter:
- Receptor systems define what physical energy the brain can receive (Property 1).
- Cortical regions and networks define where and how that energy is processed (Properties 2—3).
- Neurochemical systems modulate the processing mode (Property 4).
- The RAS sets the global activation level (Property 5).
- Inhibition sculpts precise patterns from diffuse activation (Property 6).
- Hierarchical processing builds complex representations from simple features (Property 7).
- The body connection contributes physiological state (Property 8).
- Plasticity ensures the brain adapts with repeated exposure (Property 9).
- Multiple membership, no central controller, integration, and always-on operation define the architectural constraints within which all of this operates (Properties 10—13).
Remove any one, and predictions go wrong in a specific way. Remove receptors, you lose an input channel. Remove hierarchical processing, you cannot explain why V1 activation and FFA activation have different experiential implications. Remove the always-on property, you assume a blank-state viewer who does not exist.
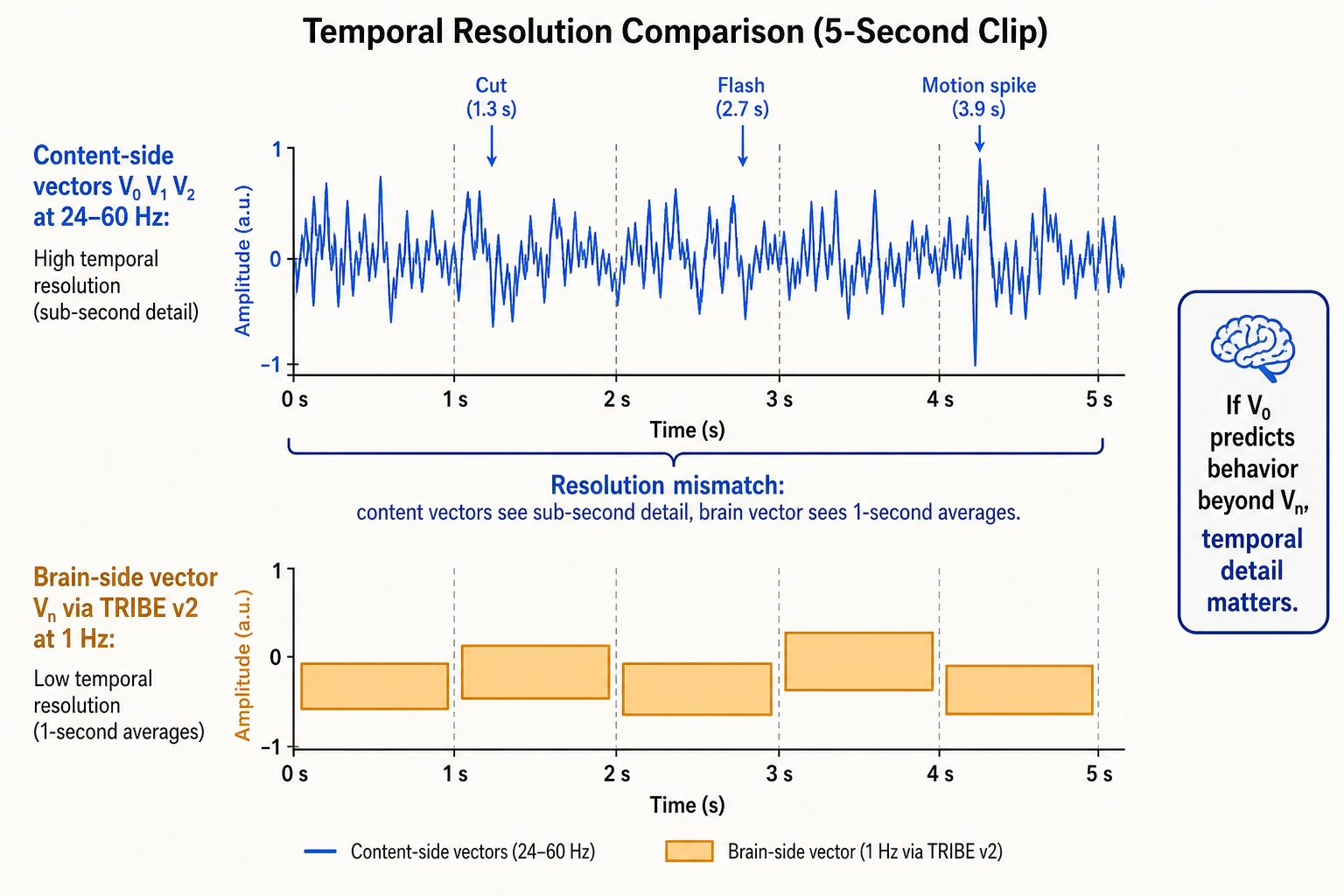
The resolution mismatch
Khozai’s vector families do not all operate at the same temporal resolution, and this matters.
| Vector family | Source | Temporal resolution | What it captures |
|---|---|---|---|
| V₀ (physical properties) | Content file, physics | 24—60 Hz (frame-by-frame) | Sub-second content dynamics with full precision |
| V₁ (temporal patterns) | Derived from V₀ | 24—60 Hz | Cuts, flashes, motion acceleration at frame level |
| V₂ (second-order patterns) | Derived from V₀ and V₁ | 24—60 Hz | How visual complexity accelerates across multi-second windows |
| Vc (cognitive dimensions) | Content file, LLM analysis | Per scene/second/shot | Semantic interpretation, not bound by brain-imaging |
| Vₙ (brain prediction) | TRIBE v2 | 1 Hz | Whole-brain activation, one prediction per second |
The content-side vectors (V₀, V₁, V₂, Vc) are computed directly from the content file. They are not limited by any brain-imaging technology. Vₙ is the only vector limited to 1 Hz, because TRIBE v2 learned from fMRI data, and fMRI detects blood-flow changes that lag 1—2 seconds behind neural activity. Everything faster is blurred into a single-second average.
The result is a resolution mismatch: the content-side vectors can detect a 0.3-second visual spike, but the brain-side vector can only report what the brain did across that entire second.
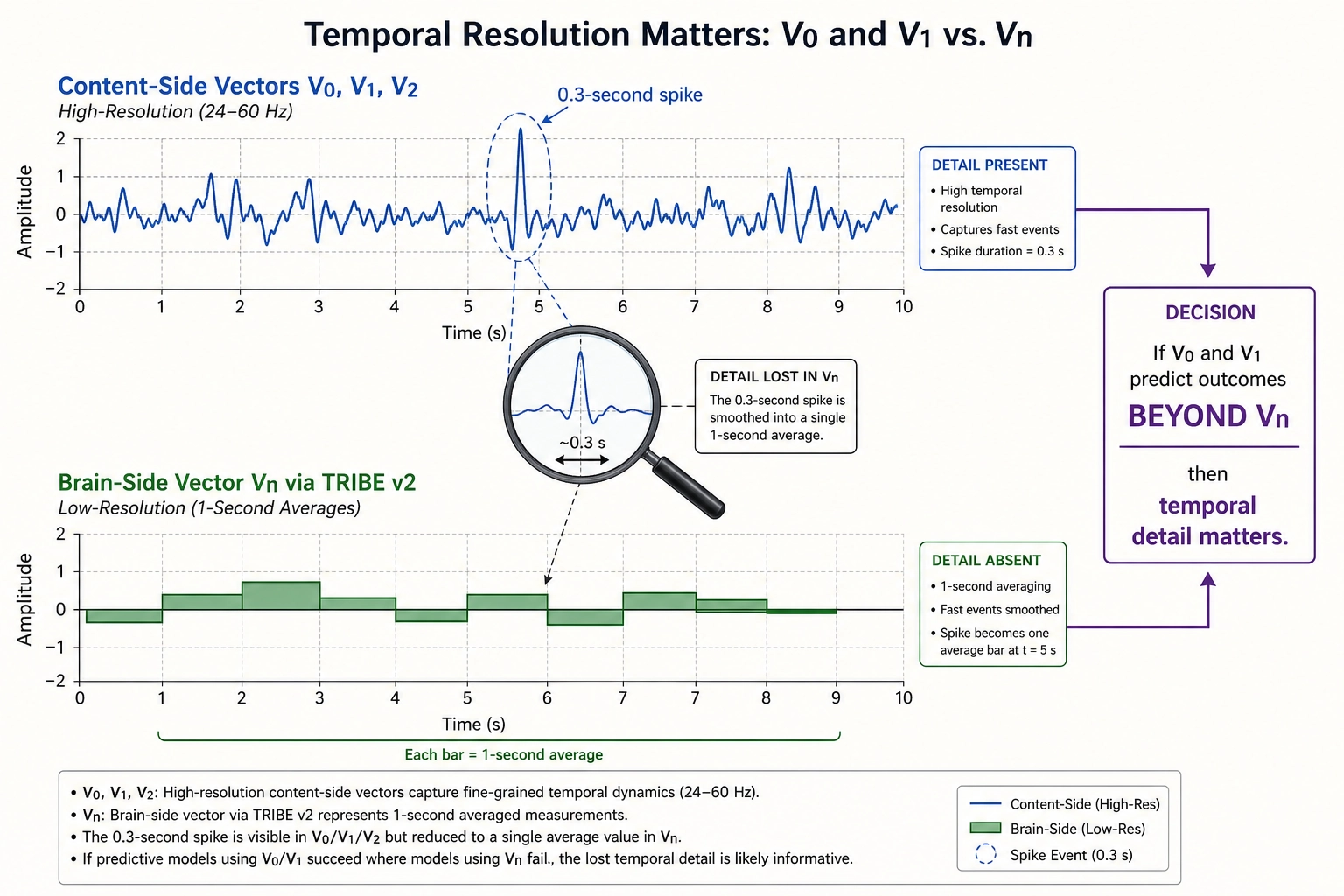
Whether this mismatch matters for Khozai’s predictions is an empirical question. The content-side vectors and the brain-side vector can check each other.
Here is the key test: if sub-second content features from V₀ and V₁ predict behavioral outcomes (views, retention, shares) BEYOND what Vₙ already explains, that is direct evidence that temporal detail matters and that Vₙ is missing something. If the answer is yes, the brain’s rapid rhythmic shifts (the alpha-to-gamma-to-theta transitions described in Section 7) likely matter for engagement, and a future complementary model (e.g., an EEG-based encoding model, if one is ever built) would close the gap. If the answer is no, Vₙ at 1 Hz already captures everything behaviorally relevant. Either way, Khozai knows. The multiple vector families are not redundant, they are the system’s way of auditing itself.
Conclusion
This chapter described the brain’s hardware, the physical machinery that Khozai’s predictions depend on.
The brain receives physical energy through ten receptor systems (Property 1), of which video content uses two: vision and hearing. It processes that energy across ~360 cortical regions organized into 7 functional networks (Properties 2—3), modulated by six neurochemical systems that set the brain’s mode, how motivated, how alert, how emotionally reactive the viewer is (Property 4). A brainstem alertness switch determines whether any processing happens at all (Property 5). Inhibition sculpts precise activation patterns from diffuse activity (Property 6). Hierarchical processing builds complex representations, faces, scenes, narratives, from simple features like edges and colors (Property 7). The body feeds its physiological state back into the brain (Property 8), and plasticity rewires connections with every exposure (Property 9). Four architectural principles, multiple membership, no central controller, integration, and always-on operation (Properties 10—13), define the constraints within which all of this operates.
Khozai consumes predictions about this entire architecture through TRIBE v2, the brain encoding model that takes a video file as input and predicts whole-brain activation, cortical predictions at higher confidence, subcortical predictions at lower confidence. The confidence gap is inherited from fMRI training data quality, not from a judgment about which structures matter. Every structure described in this chapter is in the prediction set.
Three properties make the approach viable: the architecture is finite (catalogueable, therefore measurable), hierarchically organized (decomposable, therefore analyzable at multiple resolutions), and specific (each structure maps to a specific aspect of experience, therefore traceable). Without any one of these, the chain from content to brain to experience to behavior would break.
One constraint carries forward: Khozai’s content-side vectors (V₀, V₁, V₂) operate at 24—60 Hz, but its brain-side vector (Vₙ) operates at 1 Hz. Whether this resolution mismatch matters is an empirical question the system can answer by checking whether sub-second content features predict behavioral outcomes beyond what Vₙ already explains.
This is the hardware. Chapter 4 identifies the experiential dimensions this hardware produces, what the viewer actually perceives, feels, and wants when the brain’s machinery processes a piece of content.
Bibliography
Sources are numbered in order of first appearance. TYPE tags indicate evidence strength.
[1] Coste, B. et al. “Piezo1 and Piezo2 Are Essential Components of Distinct Mechanically Activated Cation Channels.” Science, 330(6000):55-60, 2010. [PEER-REVIEWED] - Used in: Section 1 (Piezo channel discovery confirmed molecular mechanism of mechanoreception, did not add an eleventh receptor system)
[2] Brodmann, K. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues. Leipzig: Johann Ambrosius Barth, 1909. [PEER-REVIEWED] - Used in: Section 2.1 (original 52-area cytoarchitectonic atlas of the cortex)
[3] Glasser, M. F. et al. “A Multi-modal Parcellation of Human Cerebral Cortex.” Nature, 536(7615):171-178, 2016. [PEER-REVIEWED] - Used in: Section 2.1 (360-region cortical atlas from Human Connectome Project); Section 7 (Glasser region referenced for within-region heterogeneity)
[4] Yeo, B. T. T. et al. “The Organization of the Human Cerebral Cortex Estimated by Intrinsic Functional Connectivity.” Journal of Neurophysiology, 106(3):1125-1165, 2011. [PEER-REVIEWED] - Used in: Section 2.1 (7- and 17-network parcellation from 1,000 subjects)
[5] Berridge, K. C. & Robinson, T. E. “What Is the Role of Dopamine in Reward: Hedonic Impact, Reward Learning, or Incentive Salience?” Brain Research Reviews, 28(3):309-369, 1998. [PEER-REVIEWED] - Used in: Section 3 (double dissociation between wanting and liking - dopamine depletion eliminates wanting while leaving liking intact)
[6] Aston-Jones, G. & Cohen, J. D. “An Integrative Theory of Locus Coeruleus-Norepinephrine Function: Adaptive Gain and Optimal Performance.” Annual Review of Neuroscience, 28:403-450, 2005. [PEER-REVIEWED] - Used in: Section 3 (phasic vs tonic modes of the locus coeruleus - focused exploitation vs broad exploration)
[7] Moruzzi, G. & Magoun, H. W. “Brain Stem Reticular Formation and Activation of the EEG.” Electroencephalography and Clinical Neurophysiology, 1(4):455-473, 1949. [PEER-REVIEWED] - Used in: Section 4.1 (reticular activating system - stimulation produces waking, lesions produce coma)
[8] Russell, J. A. “Core Affect and the Psychological Construction of Emotion.” Psychological Review, 110(1):145-172, 2003. [PEER-REVIEWED] - Used in: Section 4.1 (two-dimensional core affect model - pleasure/displeasure and activation/arousal)
[9] Friston, K. “The Free-Energy Principle: A Unified Brain Theory?” Nature Reviews Neuroscience, 11(2):127-138, 2010. [PEER-REVIEWED] - Used in: Section 4.3 (predictive processing framework - higher cortical levels generate predictions, lower levels propagate prediction errors)
[10] Berlyne, D. E. Conflict, Arousal, and Curiosity. New York: McGraw-Hill, 1960. [PEER-REVIEWED] - Used in: Section 4.3 (inverted-U relationship between stimulus novelty and interest)
[11] Craig, A. D. “How Do You Feel? Interoception: The Sense of the Physiological Condition of the Body.” Nature Reviews Neuroscience, 3(8):655-666, 2002. [PEER-REVIEWED] - Used in: Section 5.1 (interoceptive pathway from body through brainstem to insular cortex)
[12] Damásio, A. R. Descartes’ Error: Emotion, Reason, and the Human Brain. New York: Putnam, 1994. [PEER-REVIEWED] - Used in: Section 5.1 (somatic marker hypothesis - body-state signals bias decision-making)
[13] Hebb, D. O. The Organization of Behavior: A Neuropsychological Theory. New York: Wiley, 1949. [PEER-REVIEWED] - Used in: Section 5.2 (Hebbian plasticity - neurons that fire together wire together)
[14] Zihl, J., von Cramon, D. & Mai, N. “Selective Disturbance of Movement Vision After Bilateral Brain Damage.” Brain, 106(2):313-340, 1983. [PEER-REVIEWED] - Used in: Section 7 (akinetopsia - loss of motion perception from bilateral MT/V5 lesion)
[15] Keuken, M.C., Isaacs, B.R., Trampel, R., van der Zwaag, W. & Forstmann, B.U. “Visualizing the Human Subcortex Using Ultra-high Field Magnetic Resonance Imaging.” Brain Topography, 31:513-545, 2018. [REVIEW] - Used in: Section 2.2 (subcortical fMRI signal-to-noise ratio substantially lower due to distance from coil elements, g-factor penalties, and unfavorable receive profiles)
[16] Gold, B.P., Pearce, M.T., Mas-Herrero, E., Dagher, A. & Zatorre, R.J. “Predictability and Uncertainty in the Pleasure of Music: A Reward for Learning?” Journal of Neuroscience, 39(47):9397-9409, 2019. [PEER-REVIEWED] - Used in: Section 4.3 (inverted-U relationship between prediction error and pleasure, behavioral evidence)
[17] Cheung, V.K.M., Harrison, P.M.C., Meyer, L., Pearce, M.T., Haynes, J.-D. & Koelsch, S. “Uncertainty and Surprise Jointly Predict Musical Pleasure and Amygdala, Hippocampus, and Auditory Cortex Activity.” Current Biology, 29(23):4084-4092, 2019. [PEER-REVIEWED] - Used in: Section 4.3 (uncertainty-surprise interaction predicting engagement, fMRI evidence for inverted-U)
[18] Fries, P. “Rhythms for Cognition: Communication Through Coherence.” Neuron, 88(1):220-235, 2015. [REVIEW] - Used in: Section 7 (alpha-to-gamma transition in visual cortex during attentional shifts)